Микроорганизмами (микробами) называют одноклеточные организмы размером менее 0,1 мм, которые невозможно увидеть невооруженным глазом. К ним относятся бактерии, микроводоросли, некоторые низшие мицелиальные грибы, дрожжи, простейшие (рис. 1). Их изучением занимается микробиология.
Рис. 1. Объекты микробиологии.
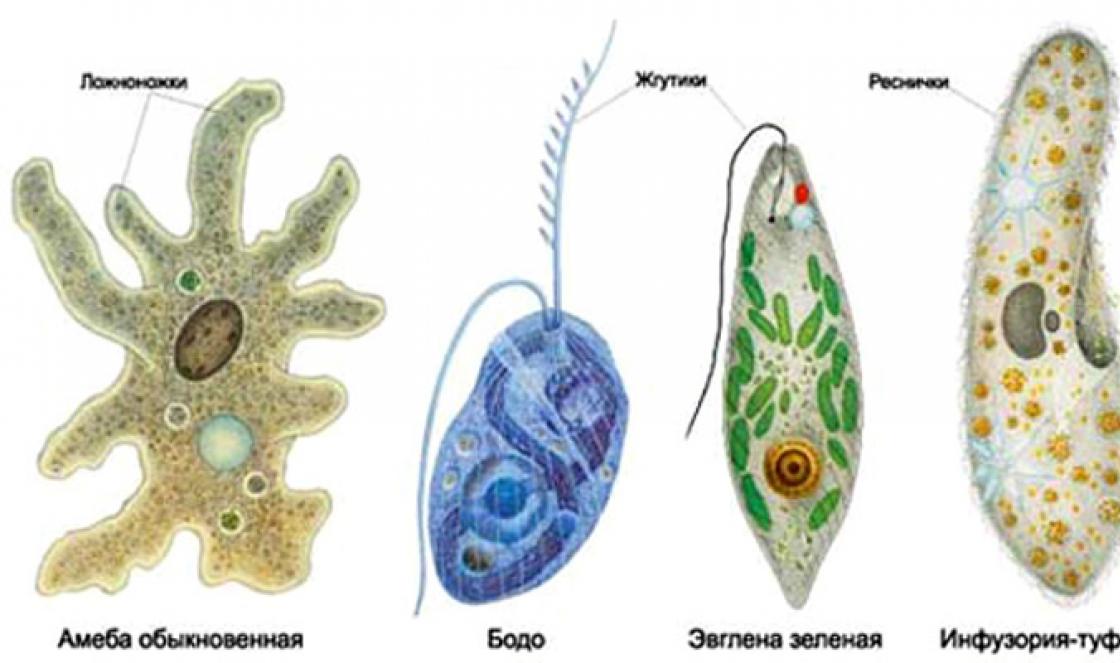
На рис. 2. можно увидеть некоторых представителей одноклеточных простейших. Иногда к объектам данной науки относят самые примитивные организмы на Земле — вирусы, не имеющие клеточную структуру и представляющие собой комплексы из нуклеиновых кислот (генетического материала) и белка. Чаще их выделяют в совершенно отдельную область исследования (Вирусологию), так как микробиология скорее направлена на изучение микроскопических одноклеточных.

Рис. 2. Отдельные представители одноклеточных эукариот (простейших).
Такие науки, как альгология и микология, изучающие водоросли и грибы, соответственно, являются отдельными дисциплинами, перекрывающимися с микробиологией в случае исследования микроскопических живых объектов. Бактериология является истинным разделом микробиологии. Данная наука занимается изучением исключительно прокариотных микроорганизмов (рис. 3).

Рис. 3. Схема прокариотической клетки.
В отличие от эукариот, к которым относятся все многоклеточные организмы, а также простейшие, микроскопические водоросли и грибы, у прокариот отсутствует оформленное ядро, содержащее генетический материал и настоящие органоиды (постоянные специализированные структуры клетки) .
К прокариотам относятся истинные бактерии и археи, по современной классификации обозначенные, как домены (надцарства) Archaea и Eubacteria (рис. 4).

Рис. 4. Домены современной биологической классификации.
Особенности строения бактерий
Бактерии являются важным звеном в круговороте веществ в природе, разлагают растительные и животные остатки, очищают загрязненные органикой водоемы, модифицируют неорганические соединения. Без них не могла бы существовать жизнь на земле. Данные микроорганизмы распространены везде, в почве, воде, воздухе, организмах животных и растений.
Бактерии различаются по следующим морфологическим особенностям:
- Форма клеток (округлые, палочковидные, нитчатые, извитые, спиралевидные, а также различные переходные варианты и звездообразная конфигурация).
- Наличие приспособлений для движения (неподвижные, жгутиковые, за счет выделения слизи).
- Сочленение клеток друг с другом (изолированные, сцепленные в виде пар, гранул, ветвящиеся формы).
Среди структур, образуемых округлыми бактериями (кокками) выделяют клетки, находящиеся в паре после деления и затем распадающиеся на одиночные образования (микрококки) или остающиеся все время вместе (диплококки). Квадратичную структуру из четырех клеток образуют тетракокки, цепочку – стрептококки, гранулу из 8-64 единиц – сарцины, грозди – стафилококки.
Палочковидные бактерии представлены многообразием форм вследствие большой изменчивости длинны (0,1-15 мкм) и толщины (0,1-2 мкм) клетки. Форма последних также зависит от способности бактерий к образованию спор – структур с толстой оболочкой, позволяющей переживать микроорганизмам неблагоприятные условия. Клетки с такой способностью называет бациллами, а не обладающие такими свойствами просто палочковидными бактериями.
Особыми видоизменениями палочковидных бактерий являются нитчатые (вытянутые) формы, цепочки и ветвящиеся структуры. Последнюю образуют актиномицеты на определенной стадии развития. «Кривые» палочки называют извитыми бактериями, среди которых выделяют вибрионы; спириллы, имеющие два изгиба (15-20 мкм); спирохеты, напоминающие волнистые линии. Их длины клеток 1-3, 15-20 и 20-30 мкм, соответственно. На рис. 5 и 6 представлены основные морфологические формы бактерий, а также типы расположения споры в клетке.

Рис. 5. Основные формы бактерий.

Рис. 6. Бактерии по типу расположения споры в клетке. 1, 4 – в центре; 2, 3, 5 – концевое расположение; 6 – с боку.
Основные клеточные структуры бактерий: нуклеоид (генетический материал), предназначенные для синтеза белка рибосомы, цитоплазматическая мембрана (часть оболочки клетки), которая у многих представителей дополнительно сверху защищена , капсулой и слизистым чехлом (рис. 7).

Рис. 7. Схема бактериальной клетки.
По классификации бактерий выделяют более 20 типов. Например, экстремально термофильные (любители высоких температур) Aquificae, анаэробные палочковидные бактерии Bacteroidetes. Однако наиболее доминантным типом, включающим в себя многообразных представителей, является Actinobacteria. К нему относятся бифидобактерии, лактобациллы, актиномицеты. Уникальность последних заключается в способности формировать мицелий на определенной стадии развития.
В простонародье это называется грибница. Действительно, разветвления клеток актиномицет напоминают гифы грибов. Несмотря на такую особенность, актиномицеты относят к бактериям, так как они являются прокариотами. Естественно их клетки по особенностям структуры менее сходны с грибами.
Актиномицеты (рис. 8) являются медленно растущими бактериями, поэтому не имеют возможности конкурировать за легкодоступные субстраты. Они способны разлагать вещества, которые другие микроорганизмы не могут использовать в качестве источника углерода, в частности углеводороды нефти. Поэтому актиномицеты интенсивно исследуются в области биотехнологии.
Некоторые представители концентрируется в зонах нефтяных месторождений, и создают специальный бактериальный фильтр, препятствующий проникновению углеводородов в атмосферу. Актиномицеты являются активными продуцентами практически ценных соединений: витаминов, жирных кислот, антибиотиков.

Рис. 8. Представитель актиномицет Nocardia.
Грибы в микробиологии
Объектом микробиологии являются только низшие плесневые грибы (ризопус, мукор, в частности). Как все грибы они не способны синтезировать вещества сами и нуждаются в питательной среде. Мицелий у низших представителей данного царства примитивен, не разделен перегородками. Особую нишу в микробиологических исследованиях занимают дрожжи (рис. 9), отличающиеся отсутствием мицелия.

Рис. 9. Формы колоний дрожжевых культур на питательной среде.
В настоящее время об их полезных свойствах собраны многочисленные знания. Однако дрожжи продолжают исследоваться на способность синтезировать практически ценные органические соединения и активно применяются в качестве модельных организмов при проведении генетических экспериментов. С древних времен дрожжи использовались в бродильных процессах. Метаболизм у разных представителей отличается. Поэтому для какого-то конкретного процесса одни дрожжи больше подходят, чем другие.
Например, Saccharomyces beticus, более устойчивые к высоким концентрациям спирта, используются для создания крепких вин (до 24%). В то время как, дрожжи S. cerevisiae способны вырабатывать более низкие концентрации этанола. По направлениям их применения дрожжи классифицируются на кормовые, пекарские, пивные, спиртовые, винные.
Болезнетворные микроорганизмы
Болезнетворные или патогенные микроорганизмы встречаются повсеместно. Наряду с широко известными вирусами: гриппом, гепатитом, корью, ВИЧ и прочее опасными микроорганизмами являются риккетсии, а также стрепто- и стафилококки, вызывающие заражение крови. Среди палочковидных бактерий много возбудителей заболеваний. Например, дифтерия, туберкулез, брюшной тиф, (рис. 10). Немало опасных для человека представителей микроорганизмов встречается среди простейших, в частности малярийный плазмодий, токсоплазма, лейшмания, лямблия, трихомонада, патогенные амебы.

Рис. 10. Фотография бактерии Bacillus anthracis, вызывающей сибирскую язву.
Многие актиномицеты не опасны для человека и животных. Однако немало патогенных представителей встречается среди микобактерий, вызывающих туберкулез, проказу (лепру). Некоторые актиномицеты инициируют такое заболевание, как актиномикоз, сопровождающийся образованием гранулем, иногда повышением температуры тела. Отдельные виды плесневых грибов способны вырабатывать токсические для человека вещества – микотоксины. Например, некоторые представители рода Aspergillus, Fusarium. Патогенные грибы вызывают группу заболеваний, называемых микозами. Так, кандидоз или, попросту говоря, молочницу вызывают дрожжеподобные грибы (рис. 11). Они всегда содержаться в организме человека, но активизируются только при ослаблении иммунитета.

Рис. 11. Гриб Candida – возбудитель молочницы.
Грибы могут вызывать разнообразные поражения кожи, в частности всевозможные виды лишая, кроме опоясывающего (герпеса), который вызывается вирусом. Дрожжи Malassezia – постоянные обитатели кожи человека при спаде активности иммунной системы могут вызвать . Не стоит сразу бежать мыть руки. Дрожжи и условно патогенные бактерии при хорошем здоровье выполняют важную функцию, препятствуют развитию болезнетворных микроорганизмов.
Вирусы как объект микробиологии
Вирусы – самые примитивные организмы на земле. В свободном состоянии в них не происходят никакие обменные процессы. Только при попадании в клетку-хозяина вирусы начинают размножаться. У всех живых организмов носителем генетического материала является дезоксирибонуклеиновая кислота (ДНК). Только среди вирусов встречаются представители с генетической последовательностью типа рибонуклеиновой кислоты (РНК).
Часто вирусы не относят к истинно живым организмам.
Морфология вирусов очень разнообразна (рис. 12). Обычно их диаметральные размеры колеблются в пределах 20-300 нм.

Рис. 12. Разнообразие вирусных частиц.
Отдельные представители достигают в длину 1-1,5 мкм. Структура вируса заключается в окружении генетического материала специальным белковым каркасом (капсидом), отличающимся разнообразием форм (спиральный, икосаэдрический, шарообразный). Некоторые вирусы сверху имеют еще оболочку, сформированную из мембраны клетки-хозяина (суперкапсид). Например, (рис. 13) известен как возбудитель заболевания, которое носит название (СПИД). Он содержит в качестве генетического материала РНК, поражает определенный тип клеток иммунной системы (т-лимфоциты хелперы).

Рис. 13. Строение вируса иммунодефицита человека.
Питание бактерий.
Питание.
Пассивная диффузия
Облегчённая диффузия
Активный транспорт
В первом случае молекула питательного вещества образует комплекс с белком периплазматического пространства, который взаимодействует со специфической пермеазой цитоплазматической мембраны. После энергозависимого проникновения через цитоплазматическую мембрану комплекс «субстрат – белок периплазмы – пермеаза» диссоциирует с освобождением молекулы субстрата.
При активном транспорте с химической модификацией переносимого вещества цепь событий включает: (1) фосфорилирование мембранного фермента-2 со стороны цитоплазмы фосфоенолпируватом; (2) связывание на поверхности цитоплазматической мембраны фосфорилированным ферментом-2 молекулы субстрата; (3) энергозависимый транспорт молекулы субстрата в цитоплазму; (4) перенос фосфатной группы на молекулу субстрата; (5) диссоциация комплекса «субстрат – фермент» в цитоплазме. За счёт фосфорилирования молекулы субстрата аккумулируются в цитоплазме клеток и не способны выйти из них.
Классификация бактерий по типу питания.
По способу поступления питательных веществ бактерии подразделяются на голофиты и голозои . Бактерии-голофиты (от греч. holos – полноценный и phyticos – относящийся к растениям) неспособны выделять в окружающую среду ферменты, расщепляющие субстраты, вследствие чего потребляют питательные вещества исключительно в растворённом, молекулярном виде. Бактерии-голозои (от греч. holos – полноценный и zoikos – относящийся к животным), напротив, имеют комплекс экзоферментов, которые обеспечивают внешнее питание – расщепление субстратов до молекул вне бактериальной клетки. После этого молекулы питательных веществ поступают внутрь бактерий-голозоев.
По источнику углерода среди бактерий выделяют автотрофы и гетеротрофы . Автотрофы (от греч. autos – сам, trophe – пища) в качестве источника углерода используют углекислый газ (СО 2), из которого синтезируют все углеродсодержащие вещества. Для гетеротрофов (от греч. geteros – другой и trophe – пища) источником углерода являются различные органические вещества в молекулярной форме (углеводы, многоатомные спирты, аминокислоты, жирные кислоты). Наибольшая степень гетеротрофности присуща прокариотам, которые могут жить только внутри других живых клеток (например, риккетсии и хламидии).
По источнику азота прокариоты подразделяются на 3 группы: 1) азотфиксирующие бактерии (усваивают молекулярный азот из атмосферного воздуха); 2) бактерии, потребляющие неорганический азот из солей аммония, нитритов или нитратов; 3) бактерии, которые ассимилируют азот, содержащийся в органических соединениях (аминокислоты, пурины, пиримидины и др.).
По источнику энергии бактерии делят на фототрофы и хемотрофы . Бактерии-фототрофы , как и растения, способны использовать солнечную энергию. Фототрофные прокариоты заболеваний у человека не вызывают. Бактерии-хемотрофы получают энергию при окислительно-восстановительных реакциях.
По природе доноров электронов литотрофы (от греч. litos – камень) и органотрофы . У литотрофов (хемолитотрофов ) в качестве доноров электронов выступают неорганические вещества (Н 2 , Н 2 S, NH 3 , сера, CO, Fe 2+ и др.). Донорами электронов у органотрофов (хемоорганотрофов ) являются органические соединения – углеводы, аминокислоты и др.
Большинство патогенных для человека бактерий обладает хемоорганотрофным (хемогетеротрофным) типом питания; реже встречается хемолитотрофный (хемоавтотрофный) тип.
По способности синтезировать органические соединения бактерии-хемотрофы подразделяются на прототрофы, ауксотрофы и гипотрофы . Бактерии-прототрофы синтезируют из глюкозы и солей аммония все необходимые органические вещества. Бактерии называются ауксотрофами , если они неспособны синтезировать какое-либо органическое вещество из указанных соединений. Крайняя степень утраты метаболической активности называется гипотрофией. Гипотрофные бактерии обеспечивают свою жизнедеятельность, реорганизуя структуры или метаболиты хозяина.
Кроме углерода и азота, для полноценной жизнедеятельности бактериям необходимы сера, фосфор, ионы металлов. Источниками серы являются аминокислоты (цистеин, метионин), витамины, кофакторы (биотин, липоевая кислота и др.), сульфаты. Источниками фосфора служат нуклеиновые кислоты, фосфолипиды, фосфаты. В достаточно высоких концентрациях бактериям нужны магний, калий, кальций, железо; в значительно меньших – цинк, марганец, натрий, молибден, медь, никель, кобальт.
Факторы роста – это вещества, которые бактерии самостоятельно синтезировать не могут, но крайне в них нуждаются. В качестве факторов роста могут выступать аминокислоты, азотистые основания, витамины, жирные кислоты, железопорфирины и другие соединения. Для создания оптимальных условий жизнедеятельности бактерий факторы роста должны быть добавлены в питательные среды.
Метаболизм, превращение энергии
А) Конструктивный метаболизм.
Обязательной фазой питания бактерий является усвоение питательных веществ, то есть включение их в изменённом или модифицированном виде в синтетические реакции по воспроизведению клеточных компонентов и структур.
Белковый обмен у бактерий может протекать в 3 фазы: первичный распад белка, вторичный распад и синтез белка. Первичный распад белковых молекул до пептонов осуществляют экзоферменты – экзопротеазы, выделяемые бактериями в окружающую среду. Вторичный распад происходит под действием эндоферментов (эндопротеаз), которые имеют все бактерии. Этот процесс протекает внутри бактериальной клетки и заключается в расщеплении пептидов до составляющих их аминокислот. Последние могут быть использованы в неизменённом виде или быть подвергнуты химическим преобразованиям (дезаминирование, декарбоксилирование и др.), в результате которых появляются аммиак, индол, сероводород, кетокислоты, спирт, углекислый газ и др. Обнаружение указанных соединений имеет в бактериологии диагностическое значение.
Наряду с реакциями расщепления белков, происходят реакции их синтеза. Одни бактерии образуют белки из готовых аминокислот, полученных в результате внешнего питания, другие бактерии самостоятельно синтезируют аминокислоты из простых соединений, содержащих азот и углерод. Синтез аминокислот может осуществляться в реакциях аминирования, переаминирования, амидирования, карбоксилирования. Большинство прокариот способны синтезировать все аминокислоты, входящие в состав клеточных белков. Особенностью биосинтеза аминокислот является использование общих биосинтетических путей: цикл трикарбоновых кислот, гликолиз, окислительный пентозо-фосфатный путь и др. Основным исходным соединением для синтеза аминокислот является пируват и фумарат.
Углеводный обмен у автотрофов и гетеротрофов имеет отличия (схема 1). Бактерии-автотрофы все необходимые углеводы синтезируют из углекислого газа. Сырьём для образования углеводов у бактерий-гетеротрофов могут служить: (1) одно-, двух- и трёхуглеродные соединения; и (2) полисахариды (крахмал, гликоген, целлюлоза). Для расщепления последних многие бактерии-гетеротрофы имеют экзоферменты (амилаза, пектиназа и др.), которые проводят гидролиз полисахаридов до образования глюкозы, мальтозы, фруктозы и пр.
У бактерий-автотрофов в цикле Кальвина из углекислого газа образуется рибулозофосфат-фосфорно-глицериновая кислота, которая включается в реакции гликолиза, идущие в обратном направлении. Конечным продуктом обратного синтеза является глюкоза.
Бактерии-гетеротрофы образуют глюкозу из одно-, двух- и трёхуглеродных соединений, также включая их в реакции обратного гликолиза. Ввиду того, что некоторые реакции гликолиза необратимы, у гетеротрофов сформировались специальные ферментативные реакции, позволяющие обходить необратимые реакции катаболического пути.
При расщеплении бактериями-гетеротрофами полисахаридов образующиеся дисахариды поступают внутрь клеток и под влиянием мальтозы, сахарозы, лактозы подвергаются гидролизу и распаду на моносахара, которые затем сбраживаются либо включаются в реакции взаимопревращения сахаров.
Липидный обмен . Исходными материалами для образования липидов у бактерий могут служить как экзогенные липиды, так и амфиболиты межуточного обмена. Экзогенные липиды подвергаются действию бактериальных липаз и других липолитических ферментов. Многие виды бактерий усваивают глицерин, который служит источником пластического материала и энергии. Эндогенными источниками для синтеза липидов могут быть ацетилкоэнзим А, пропионил-АПБ, малонил-АПБ (АПБ – ацетилпереносящий белок), фосфодиоксиацетон и др.
Исходным субстратом для синтеза жирных кислот с чётным числом углеродных атомов служит ацетилкоэнзим А, для жирных кислот с нечётным числом углеродных атомов – пропионил-АПБ и малонил-АПБ. Образование двойных связей в молекуле кислоты у аэробных прокариот происходит при участии молекулярного кислорода и фермента десатуразы. У анаэробных прокариот двойные связи вводятся на ранних этапах синтеза в результате реакции дегидратации. Исходным субстратом для синтеза фосфолипидов служит фосфодиоксиацетон (промежуточное соединение гликолитического пути), восстановление которого приводит к образованию 3-фосфороглицерина. К последнему затем присоединяются 2 остатка жирных кислот в виде комплекса с АПБ. Продуктом реакции является фосфатидная кислота, активирование которой с помощью ЦТФ и последующее присоединение к фосфатной группе серина, инозита, глицерина или другого соединения приводят к синтезу соответствующих фосфолипидов.
Ауксотрофные и гипотрофные по жирным кислотам микроорганизмы (например, микоплазмы) получают их в готовом виде из клеток хозяина или питательной среды.
Мононуклеотидный обмен . Пуриновые и пиримидиновые мононуклеотиды являются важнейшими компонентами ДНК и РНК. Многие прокариоты способны как использовать содержащиеся в питательной среде готовые пуриновые и пиримидиновые основания, их нуклеозиды и нуклеотиды, так и синтезировать их из низкомолекулярных веществ. Бактерии располагают ферментами, катализирующими следующие этапы взаимопревращений экзогенных пуриновых и пиримидиновых производных: азотистое основание – нуклеозид – нуклеотид (моно- – ди- – трифосфат).
Синтез пуриновых и пиримидиновых мононуклеотидов de novo осуществляется независимыми путями. При синтезе пуриновых нуклеотидов в результате последовательных ферментативных реакций образуется инозиновая кислота, из которой путём химических модификаций пуринового кольца синтезируются адениловая (АМФ) и гуаниловая (ГМФ) кислоты. Синтез пиримидиновых нуклеотидов начинается с образования оротидиловой кислоты, декарбоксилирование которой даёт уридиловую кислоту (УМФ). Из последней образуется УТФ, ацилирование которого приводит к возникновению ЦТФ.
Дезоксирибонуклеотиды образуются в результате восстановления соответствующих рибонуклеотидов на уровне дифосфатов или трифосфатов. Синтез специфического для ДНК нуклеотида – тимидиловой кислоты происходит путём ферментативного метилирования дезоксиуридиловой кислоты.
Ионный обмен . Минеральные соединения – ионы, NH 3 + , К + , Mg 2+ , Fe 2+ , SO 4 2- , PO 4 3- и другие бактерии получают из окружающей среды как в свободном, так и в связанном с другими органическими веществами состоянии. Катионы и анионы транспортируются в бактериальную клетку различными способами, описанными в § 3. На скорость проникновения ионов в бактериальную клетку влияют рН среды и физиологическая активность самих микроорганизмов.
Б) Дыхание бактерий (энергетический метаболизм).
Все процессы жизнедеятельности энергозависимы, поэтому добывание энергии является крайне важной стороной метаболизма прокариот. Они получают энергию при анаэробном и аэробном дыхании.
Дыхание , или биологическое окисление – это катаболический процесс переноса электронов от вещества-донора на вещество-акцептор, сопровождающийся накоплением энергии в макроэргических соединениях . Дыхание осуществляется в процессе катаболических реакций, в результате которых сложные органические вещества, расщепляясь, отдают энергию и превращаются в простые соединения. Аккумулированная в макроэргических веществах (АТФ, ГТФ, УТФ и др.) энергия используется в анаболических реакциях.
По способу дыхания микроорганизмы подразделяются на облигатные (строгие) аэробы, облигатные анаэробы и факультативные анаэробы .
Облигатные аэробы нуждаются в свободном кислороде. Донорами электронов у патогенных для человека аэробов-хемоорганотрофов являются органические соединения (углеводы, жиры, белки), акцептором электронов – молекулярный кислород. Запасание энергии в виде АТФ у аэробов-хемоорганотрофов происходит при окислительном фосфорилировании доноров электронов. Аэробы обладают цитохромами (участвуют в переносе электронов), а также ферментами (каталаза, супероксиддисмутаза, пероксидаза), инактивирующими токсические кислородные радикалы, образующиеся при дыхании. Супероксиддисмутаза инактивирует наиболее токсичный метаболит – супероксидрадикал в Н 2 О 2 . Фермент каталаза превращает Н 2 О 2 в Н 2 О и О 2 .
Особую группу аэробов составляют микроаэрофильные бактерии , которые хотя и нуждаются в кислороде для получения энергии, лучше растут при повышенном содержании СО 2 , например, бактерии родов Campylobacter и Helicobacter .
Облигатные анаэробы не нуждаются в свободном кислороде, напротив, даже в малых количествах кислород оказывает на них токсическое действие. Донорами электронов у патогенных для человека анаэробов-хемоорганотрофов служат различные органические соединения (преимущественно углеводы). Акцептором электронов у анаэробов-хемоорганотрофов являются органические кислородсодержащие соединения – кислоты или кетоны, то есть акцептор электрона - связанный с органическим фрагментом кислород. Запасание энергии у этих прокариот происходит при субстратном фосфорилировании. Облигатные анаэробы, как правило, не имеют цитохромов и ферментов, инактивирующих кислородные радикалы (каталазо- и супероксидисмутазоотрицательны).
У непатогенных для человека анаэробов хемолитотрофов акцептором электронов являются неорганические кислородсодержащие соединения – нитраты, сульфаты, карбонаты.
Особую группу анаэробов составляют аэротолерантные бактерии, которые способны расти в присутствии атмосферного кислорода, но не используют его в качестве акцептора электронов (например, молочнокислые бактерии). Аэротолерантные прокариоты каталазо- и супероксиддисмутазопозитивны.
Факультативные анаэробы способны существовать как в кислородной, так и в бескислородной средах. Донорами электронов у них являются органические вещества; акцепторами электронов, в зависимости от условий среды – молекулярный или связанный в органических и неорганических соединениях кислород. Энергия факультативными анаэробами может аккумулироваться как при окислительном, так и при субстратном фосфорилировании. Как и аэробы, данная группа бактерий имеет цитохромы и ферменты антиоксидантной защиты.
Основным субстратом для получения энергии являются углеводы, которые у разных по типу дыхания хемогетеротрофных прокариот могут катаболизироваться до ацетилкоэнзима А («активированная уксусная кислота»). В качестве энергетических субстратов могут выступать липиды и белки, поскольку ацетилкоэнзим А также является одним из промежуточных продуктов их метаболизма (схема 2).
Катаболизм углеводов у хемоорганотрофных прокариот включает: (а) анаэробные процессы – гликолиз, пентозофосфатный путь и кетодезоксифосфоглюконатный путь; (б) аэробный процесс – цикл трикарбоновых кислот (цикл Кребса). Анаэробные процессы имеют место у всех прокариот, тогда как аэробный процесс характерен только для облигатных аэробов и факультативных анаэробов. В основе получения энергии анаэробными путями лежит субстратное фосфорилирование, в основе аэробного процесса – окислительное фосфорилирование.
Определение понятий.
Стерилизация, дезинфекция и антисептика являются неотъемлемыми частями современной медицинской и в особенности хирургической практики. Понимание принципов и практического применения этих методов необходимо, поскольку многие потенциально патогенные микроорганизмы способны оставаться жизнеспособными вне макроорганизма в течение длительного времени, проявлять высокую устойчивость к действию физических и химических дезинфектантов и относительно легко передаваться от одного человека к другому.
Антисептика - уничтожение или предотвращение роста патогенных или условно-патогенных микроорганизмов химическими методами. Этот термин обычно используют для обозначения наружного нанесения химического препарата на живые ткани.
Антисептик - вещество, которое угнетает рост или разрушает микроорганизм (без действия на споры бактерий). Термин является специфическим для обозначения веществ, которые используются для местного действия на живые ткани.
Асептика означает отсутствие сепсиса, но вообще этот термин используют для того, чтобы подчеркнуть отсутствие любых живых организмов. Асептические методы означают любую процедуру, предназначенную для элиминации живых организмов и предотвращения повторной контаминации ними. Современные хирургические и микробиологические методы основаны на асептических процедурах.
Биоцид - вещество, которое убивает все живые микроорганизмы, как патогенные, так и непатогенные, включая споры.
Биостат - агент, который предотвращает рост микроорганизмов, но необязательно убивает их.
Деконтаминация - удаление микроорганизмов без количественного определения. Этот термин является относительным; окончательное удаление микробов может быть осуществлено стерилизацией или дезинфекцией.
Дезинфекция - процесс, который уменьшает количество или полностью уничтожает все патогенные микроорганизмы, кроме спор.
Гермицид - вещество, которое разрушает микроорганизмы, особенно патогенные. Гермицид не разрушает споры.
Санация - метод, благодаря которому микробная контаминация уменьшается до “безопасного” уровня. Этот метод ранее использовали для “очищения” неживых объектов.
Стерилизация - использование физических факторов и (или) химических веществ для полного уничтожения или разрушения всех форм микробной жизни.
Стерилизация.
Стерилизацию определяют как разрушение или удаление (путем фильтрации) всех микроорганизмов и их спор. Стерилизацию обычно проводят с помощью тепла. Стерилизация, будучи одной из повседневных процедур в работе микробиологической лаборатории, является необходимым методом, обеспечивающим такую обработку, при которой культуры, оборудование, посуда и среды способствуют росту только необходимых микроорганизмов, тогда как другие микробы разрушаются. Различают такие виды стерилизации: прокаливание в пламени горелки, кипячение, действие текучим паром, паром под давлением в автоклаве, сухим жаром, пастеризация, тиндализация, химическая, холодная (механическая) стерилизация.
Выбор методов стерилизации.
При выборе методов стерилизации нужно учитывать следующие требования:
1. Активность: бактерицидная, спороцидная, туберкулоцидная, фунгицидная и вирусоцидная.
2. Скорость процедуры: стерилизация должна проводиться как можно более быстро.
3. Проницаемость: вещества-стерилизаторы должны проникать через упаковку и к внутренним частям инструментария.
4. Совместимость: не должны возникать изменения структуры или функции материалов, которые стерилизуют несколько раз.
5. Нетоксичность: не должно возникать угрозы для здоровья человека и состояния окружающей среды.
6. Устойчивость органического материала: эффективность стерилизации не должна снижаться в присутствия органического материала.
7. Приспособляемость: возможность использовать для больших и малых объёмов стерилизуемого материала.
8. Контроль в течение времени: цикл обработки должен легко и точно контролироваться.
9. Цена: разумная стоимость оснащения, установки и эксплуатации.
 Физические стерилизаторы
Физические стерилизаторы
Влажное тепло, которое образуется в процессе парового автоклавирования, является основным стерилизующим агентом, используемым в лабораториях клинической микробиологии. Автоклавы используют для стерилизации питательных сред, жароустойчивых материалов и обработки инфицированных отходов. Паровой стерилизатор, или автоклав, представляет собой изолированную камеру под давлением, которая использует насыщенный пар для создания высоких температур (рис. 1). Воздух удаляют из камеры замещением по массе или созданием вакуума. Наиболее часто используют автоклавы с замещением по массе. Более лёгкий пар запускают в камеру для вытеснения более тяжёлого воздуха. Кратковременная обработка паром под давлением может уничтожить бактериальные споры. Для рутинной стерилизации питательных сред и других материалов время экспозиции составляет 15 минут при 121ºС и давление - 1,5 кг на 1 квадратный сантиметр. Для инфекционных отходов время экспозиции увеличивается до 30-60 минут. Дополнительно к правильно выбранным времени и температуре, очень важным при стерилизации является прямой контакт с паром. При обработке инфекционного материала следует обеспечить максимальное проникновение пара в отходы. Такой материал необходимо обрабатывать при температуре 132ºС. Не подлежат автоклавированию антинеопластические препараты, токсичные химические вещества и радиоизотопы, которые могут не разрушиться, а также нестабильные химикаты, поскольку они под действием тепла могут испариться и распространиться по камере.
 Стерилизация сухим жаром
используется для материалов, которые невозможно стерилизовать паром в связи с возможностью повреждения или в связи с непроницаемостью материала для пара. Сухой жар менее эффективен, чем влажное тепло, и требует болеего времени экспозиции и более высоких температур. Стерилизацию сухим жаром обычно проводят в сухожаровом шкафу (рис. 2). Механизм стерилизации с помощью сухого жара является окислительным процессом. Примерами материалов, для которых используют стерилизацию сухим жаром, являются масла, порошки, острые инструменты и стеклянная посуда. Сухой жар или термическую инактивацию-стерилизацию используют как альтернативные методы обработки инфекционных отходов.
Стерилизация сухим жаром
используется для материалов, которые невозможно стерилизовать паром в связи с возможностью повреждения или в связи с непроницаемостью материала для пара. Сухой жар менее эффективен, чем влажное тепло, и требует болеего времени экспозиции и более высоких температур. Стерилизацию сухим жаром обычно проводят в сухожаровом шкафу (рис. 2). Механизм стерилизации с помощью сухого жара является окислительным процессом. Примерами материалов, для которых используют стерилизацию сухим жаром, являются масла, порошки, острые инструменты и стеклянная посуда. Сухой жар или термическую инактивацию-стерилизацию используют как альтернативные методы обработки инфекционных отходов.
Пастеризация разрушает патогенные микроорганизмы путём быстрого нагревания вещества до 71,1ºС на протяжении 15 с, что сопровождается последующим быстрым охлаждением. Пастеризация не является стерилизацией, поскольку не все микроорганизмы чувствительны к ней. Этот метод элиминировал пищевой путь передачи таких заболеваний, как туберкулез пищеварительного тракта и Q-лихорадка.
Тиндализация - это метод стерилизации прерывистым нагреванием, который может использоваться для уничтожения всех бактерий в растворах. Поскольку растущие бактерии легко гибнут при кратковременном кипячении (5 раз в течение 1 часа по 5 минут), всё, что необходимо сделать, это позволить раствору постоять на протяжении определенного времени, прежде чем тепло нарушит созревание спор с существенной потерей их устойчивости к теплу.
Фильтрация - это процесс, который используют для удаления микробов и микроскопических частей из растворов, воздуха и других газов. Наиболее часто стерилизацию путем фильтрации в лаборатории используют для обработки диагностических препаратов, питательных сред, тканевых культуральных сред, сывороток, растворов, которые содержат компоненты сыворотки. Другим общепринятым применением фильтрации является стерилизация воздуха и газов. Пластиковые или бумажные мембранные фильтры, которые различают по диаметру пор (примерно от 12 до 0,22 μм) и используют для механического разделения, служат и для сбора микробов из жидкостей для микроскопического изучения или культивирования прямо на фильтре, когда его помещают на поверхность, пропитанную питательной средой.
Ультрафиолетовое облучение является видом электромагнитной волновой радиации, которая действует на клеточную нуклеиновую кислоту. Микроорганизмы высокочувствительны к действию ультрафиолетовых лучей с длиной волны 254 нм. Ультрафиолет наиболее широко используют для уничтожения микроорганизмов, находящихся в воздухе или на каких-либо поверхностях. Другим применением является холодная стерилизация определенных химикатов и пластика для фармацевтических целей, стерилизация сыворотки для клеточных культур и дезинфекция воды. Существенным недостатком ультрафиолетового облучения в качестве стерилизатора является его неспособность к проникновению внутрь материалов.
Ионизирующее излучение в электромагнитном спектре летально действует на микроорганизмы. Этот спектр включает микроволны, γ-лучи, рентгеновские лучи и поток электронов. Летальный эффект от ионизирующего излучения возникает вследствие прямого действия на молекулу-мишень, в результате чего энергия переносится в молекулу; и вследствие косвенного действия - диффузии радикалов.
Ультразвуковая энергия с низкой частотой инактивирует микроорганизмы в водных растворах. Физический эффект обработки ультразвуком возникает вследствие кавитации. Ультразвуковые очистители и другие приборы часто используют для очистки инструментов, но не считают стерилизаторами. Однако комбинирование ультразвука с химической обработкой убивает микроорганизмы.
Химические стерилизаторы
2 % глютаровый альдегид в качестве жидкого химического стерилизатора ранее широко применяли для обработки медицинского и хирургического материала, который невозможно стерилизовать нагреванием или облучением. Глютаровый альдегид также используют при приготовлении вакцин.
Дезинфекция.
Дезинфекцию можно проводить химическими методами или кипячением. Кипячение является эффективным методом дезинфекции инструментария, например, игл и шприцев, если нет автоклава. Предварительно очищенный медицинский инструментарий следует кипятить 20 минут. Химическую дезинфекцию используют для чувствительного к действию тепла оборудования, которое может повредить высокая температура. Широко используют такие химические дезинфектанты, как компонента хлора, этиловый и изопропиловый спирт, четвертичные компоненты аммония и глютаровый альдегид.
Химические дезинфектанты.
Спирт (этиловый и изопропиловый) , растворённый в воде до концентрации 60-85 %, очень эффективен при дезинфекции. Спирты имеют бактерицидное, фунгицидное и туберкулоцидное действие, но не влияют на споры. Этиловый спирт имеет более широкий спектр вирусоцидной активности, чем изопропиловый, поэтому он более эффективно действует на липофильные и гидрофильные вирусы.
Раствор 37 % формальдеида , который называют формалином, можно использовать в качестве стерилизатора, тогда как его концентрации 3-8 % можно использовать в качестве дезинфектантов.
Фенол в чистом виде не используют в качестве дезинфектанта в связи с его токсичностью, способностью индуцировать развитие опухолей и коррозии. Дериваты фенола, в которых функциональная группа (хлор, бром, алкил, бензил, фенил, амил) замещает один из атомов водорода в ароматическом кольце, широко используют в качестве дезинфектантов. Подобное замещение уменьшает недостатки фенола. Компоненты фенола убивают микробы благодаря инактивации ферментных систем, преципитации белков и нарушению клеточной стенки и мембраны. Обычно используют концентрации 2-5 %, более низкая концентрация требует более длительной экспозиции.
Галогены. Только хлор и йод используют для дезинфекции в лабораторной практике. В связи с тем, что хлор является мощным окислителем, считают, что он убивает микробы путем окисления. Считают, что йод убивает микроорганизмы путём реакции с N-H и S-H группами аминокислот, а также с фенольной группой аминокислоты тирозина и углерод-углеродными двойными связями ненасыщенных жирных кислот. Обычная обработка включает распыление 2-5 % раствора формальдегида в присутствии пара при температуре 60-80ºС.
Антисептика.
Антисептики можно обнаружить в микробиологических лабораториях, прежде всего, в веществах, которые используют для мытья рук. В тех случаях, когда медицинский персонал оказывает неотложную помощь пациентам с использованием веществ, содержащих антибактериальные агенты, это уменьшает количество госпитальных инфекций. Наиболее распространёнными химическими соединениями, содержащимися в веществах для мытья рук, являются спирты, хлоргексидина глюконат, йодофоры, хлороксайленол и триклозан.
Традиционными методами обработки отходов и мусора являются сжигание и стерилизация паром.
Сжигание является методом выбора для обработки отходов и мусора. Этот метод делает отходы неинфекционными, а также изменяет их форму и размеры. Стерилизация является эффективным методом обработки отходов, но она не изменяет их формы. Стерилизация паром в автоклаве при 121ºС в течение минимум 15 минут уничтожает все формы микробной жизни, включая большое количество бактериальных спор. Этот тип полной стерилизации также можно провести с использованием сухого жара при температуре 160-170ºС на протяжении 2-4 часов. Однако следует убедиться, что сухой жар контактирует со стерилизуемым материалом. Поэтому бутылки, которые содержат жидкость, должны быть неплотно закрыты пробками или ватными тампонами для того, чтобы пар и жар могли обмениваться с воздухом в бутылках. Биологически опасные контейнеры, содержащие отходы, следует плотно завязать. Простерилизованный биологически опасный материал нужно запечатать в соответствующие контейнеры с этикетками.
Стерилизация паром (в автоклаве). Инфекционный мусор считают деконтаминированным при уменьшении в 6 lg раз количества вегетативных бактерий, грибов, микобактерий и вирусов, содержащих липиды, и в 4 lg раза - бактериальных эндоспор.
Питание бактерий.
Питание. Под питанием бактериальной клетки следует понимать процесс поглощения и усвоения пластического материала и энергии в результате преобразовательных реакций . Типы питания прокариот сложны и разнообразны. Они различаются в зависимости от способа поступления питательных веществ внутрь бактериальной клетки, источников углерода и азота, способа получения энергии, природы доноров электронов.
Транспорт питательных веществ внутрь клетки может осуществляться 3 механизмами: пассивной диффузией, облегчённой диффузией и активным транспортом.
Пассивная диффузия является неспецифическим энергозависимым процессом, осуществляемым по градиенту концентрации веществ (вещество из среды с большей своей концентрацией пассивно, согласно законам осмоса, поступает в среду с меньшей концентрацией). Пассивной диффузией внутрь бактериальной клетки поступает ограниченное количество веществ, некоторые ионы, моносахара. Скорость переноса веществ при пассивной диффузии незначительна и зависит от липофильности и размеров транспортирующихся молекул.
Облегчённая диффузия представляет собой энергонезависимый транспорт веществ по градиенту концентрации при помощи ферментов пермеаз. Пермеазы – это специфические мембранные белки, способствующие прохождению веществ через цитоплазматическую мембрану. Пермеаза фиксирует на себе молекулу переносимого вещества, вместе с которым пеодолевает цитоплазматическую мембрану, после чего комплекс «вещество – пермеаза» диссоциирует. Освободившаяся пермеаза используется для проведения других молекул. У прокариотов облегчённой диффузией внутрь клетки поступает только глицерин. При этом внутриклеточная концентрация глицерина соответствует таковой вне клетки. Облегчённая диффузия наиболее характерна для микроорганизмов-эукариот.
Активный транспорт – это энергозависимый перенос веществ внутрь клетки против градиента концентрации при помощи специфических ферментов. Активным транспортом в бактериальную клетку поступает подавляющее большинство веществ (ионы, углеводы, аминокислоты, липиды и др.). Активный транспорт может осуществляться: (1) без химической модификации переносимого вещества; (2) с химической модификацией.
Какие бывают бактерии: виды бактерий, их классификация
Бактерии — это крошечные микроорганизмы, которые появились много тысячелетий назад. Увидеть микробы невооруженным глазом невозможно, но не следует забывать об их существовании. Существует огромное количество бацилл. Их классификацией, изучением, разновидностями, особенностями строения и физиологии занимается наука микробиология.
Микроорганизмы по-разному называются, в зависимости от своего рода действий и функций. Под микроскопом можно наблюдать, как эти маленькие существа взаимодействуют друг с другом. Первые микроорганизмы были довольно примитивными по форме, но и их значение ни в коем случае нельзя преуменьшать. С самого начала бациллы развивались, создавали колонии, пытались выжить в изменчивых климатических условиях. Разные вибрионы способны обмениваться аминокислотами, чтобы в результате нормально расти, развиваться.
Сегодня трудно сказать, сколько на земле есть видов этих микроорганизмов (это число превышает миллион), но самые известные и их названия знакомы практически каждому человеку. Неважно, какие бывают и как называются микробы, все они имеют одно преимущество — они живут колониями, так им намного легче адаптироваться и выживать.
Для начала давайте разберемся, какие существуют микроорганизмы. Самая простая классификация — это хорошие и плохие. Другими словами те, которые несут вред человеческому организму, становятся причиной многих болезней и те, которые приносят пользу. Далее мы поговорим детально, какие есть основные полезные бактериии дадим их описание.

Можно также классифицировать микроорганизмы соответственно их форме, характеристике. Наверное, многие помнят, что в школьных учебниках была специальная таблица с изображением разных микроорганизмов, а рядышком было значение и их роль в природе. Есть несколько типов бактерий:
- кокки — небольшие шарики, которые напоминают цепочку, так как располагаются друг за дружкой;
- палочковидные;
- спириллы, спирохеты (имеют извитую форму);
- вибрионы.
Бактерии разных форм
Мы уже упоминали, что одна из классификаций делит микробы на виды в зависимости от их форм.
Бактерии палочки тоже имеют некоторые особенности. Например, есть виды палочковидных с заостренными полюсами, с утолщенными, с закругленными или же с прямыми концами. Как правило, палочковидные микробы очень разные и всегда находятся в хаосе, они не выстраиваются цепочкой (за исключением стрептобацилл), не крепятся друг к дружке (кроме диплобацилл).
К микроорганизмам шаровидных форм микробиологи относят стрептококки, стафилококки, диплококки, гонококки. Это могут быть пары или же длинные цепочки из шариков.
Изогнутые бациллы — это спириллы, спирохеты. Они всегда активны, но не производят спор. Спириллы безопасны для людей, для животных. Отличить спириллы от спирохет можно, если обратить внимание на количество завитков, они менее извиты, имеют специальные жгутики на конечностях.
Виды болезнетворных бактерий
Например, группа микроорганизмов под названием кокки, а более детально стрептококки и стафилококки становятся причиной настоящих гнойных заболеваний (фурункулез, стрептококковая ангина).
Анаэробы прекрасно живут и развиваются без кислорода, для некоторых типов этих микроорганизмов кислород вообще становится смертельным. Аэробные микробы нуждаются в кислороде для полноценного существования.
Археи— это практически бесцветные одноклеточные организмы.
Патогенных бактерий нужно остерегаться, ведь они вызывают инфекции, грамотрицательные микроорганизмы считаются устойчивыми к антителам. Много информации есть о почвенных, гнилостных микроорганизмах, которые бывают вредными, полезными.

В общей сложности спириллы не представляют собой опасности, но некоторые виды могут вызывать содоку.
Разновидности полезных бактерий
О том, что бациллы бывают полезные и вредные, знают даже школьники. Некоторые названия люди знают на слух (стафилококк, стрептококк, чумная палочка). Это вредные существа, которые мешают не только внешней среде, но и человеку. Есть микроскопические бациллы, которые вызывают пищевые отравления.
Обязательно нужно знать полезную информацию о молочнокислых, пищевых, пробиотических микроорганизмах. Например, пробиотики, иными словами хорошие организмы, часто применяют в медицинских целях. Вы спросите: для чего? Они не позволяют вредным бактериям размножаться внутри человека, укрепляют защитные функции кишечника, хорошо влияют на иммунную систему человека.
Бифидобактерии также очень полезны для кишечника. Молочнокислые вибрионы включают в себя около 25 видов. В человеческом организме они имеются в огромных количествах, но не являются опасными. Наоборот, защищают желудочно-кишечный тракт от гнилостных и других микробов.
Говоря о хороших, нельзя не упомянуть и огромный вид стрептомицетов. Они известны тем, кто принимал левомицетин, эритромицин и подобные препараты.
Есть такие микроорганизмы, как азотобактеры. Они много лет живут в грунтах, благотворно влияют на почву, стимулируют рост растений, очищают землю от тяжелых металлов. Они незаменимы в медицине, сельском хозяйстве, медицине, пищевой промышленности.
Виды изменчивости бактерий
По своей природе микробы очень непостоянные, они быстро умирают, они могут быть спонтанными, индуцированными. Мы не будем вдаваться в подробности об изменчивости бактерий, так как эта информация больше интереснатем, кого интересует микробиология и все ее ответвления.
Виды бактерий для септиков
Жители частных домов понимают острую необходимость очищать сточные воды, а также выгребные ямы. Сегодня быстро и качественно очистить стоки можно с помощью специальных бактерий для септиков. Для человека это огромное облегчение, так как заниматься чисткой канализации—дело не из приятных.
Мы уже прояснили, где применяется биологический вид очистки стоков, а теперь поговорим о самой системе. Бактерии для септиков выращиваются в лабораториях, они убивают неприятный запах стоков, дезинфицируют дренажные колодцы, выгребные ямы, уменьшают объем сточных вод. Есть три вида бактерий, которые используются для септиков:
- аэробные;
- анаэробные;
- живые (биоактиваторы).

Очень часто люди используют комбинированные методы очистки. Строго следуйте инструкциям на препарате, следите, чтобы уровень воды способствовал нормальному выживанию бактерий. Также не забывайте использовать канализацию как минимум раз в две недели, чтобы бактериям было чем питаться, иначе они умрут. Не забывайте, что хлор из порошков и жидкостей для чистки, убивает бактерии.
Самыми популярными являются бактерии Доктор Робик, Септифос, Вэйст Трит.
Виды бактерий в моче
По идее бактерий в моче быть не должно, но после различных действий и ситуаций, крошечные микроорганизмы поселяются, где им вздумается: во влагалище, в носу, в воде и так далее. Если бактерии были обнаружены во время анализов, это означает, что человек страдает от болезней почек, мочевого пузыря или мочеточников. Есть несколько путей, по которым микроорганизмы попадают в мочу. Перед лечением очень важно исследовать и точно определить тип бактерий и способ попадания. Определить это можно при биологическом посеве мочи, когда бактерии помещают в благоприятную среду обитания. Далее проверяется реакция бактерий на различные антибиотики.
Мы желаем вам оставаться всегда здоровыми. Следите за собой, регулярно мойте руки, берегите свой организм от вредоносных бактерий!
2.1. Систематика и номенклатура микробов
Мир микробов можно разделить на клеточные и неклеточные формы. Клеточные формы микробов представлены бактериями, грибами и простейшими. Их можно называть микроорганизмами. Неклеточные формы представлены вирусами, вироидами и прионами.
Новая классификация клеточных микробов включает следующие таксономические единицы: домены, царства, типы, классы, порядки, семейства, роды, виды. В основу классификации микроорганизмов положены их генетическое родство, а также морфологические, физиологические, антигенные и молекулярнобиологические свойства.
Вирусы нередко рассматриваются не как организмы, а как автономные генетические структуры, поэтому они будут рассмотрены отдельно.
Клеточные формы микробов разделены на три домена. Домены Bacteria и Archaebacteria включают микробы с прокариотическим типом строения клетки. Представители домена Eukarya являются эукариотами. Он состоит из 4 царств:
Царства грибов (Fungi, Eumycota);
царства простейших (Protozoa);
царства Chromista (хромовики);
Микробов с неуточненным таксономическим положением (Microspora, микроспоридии).
Различия в организации прокариотической и эукариотической клеток представлены в табл. 2.1.
Таблица 2.1. Признаки прокариотической и эукариотической клетки
2.2. Классификация и морфология бактерий
Термин «бактерия» происходит от слова bacterion, что означает палочка. Бактерии относятся к прокариотам. Их разделяют на два домена: Bacteria и Archaebacteria. Бактерии, входящие в домен Archaebacteria, представляют одну из древнейших форм жизни. Они имеют особенности строения клеточной стенки (у них отсутствует пептидогликан) и рибосомальной РНК. Среди них отсутствуют возбудители инфекционных заболеваний.
Внутри домена бактерии подразделяются на следующие таксономические категории: класс, тип, порядок, семейство, род, вид. Одной из основных таксономических категорий является вид (species). Вид - это совокупность особей, имеющих единое происхождение и генотип, объединенные по близким свойствам, отличающим их от других представителей рода. Название вида соответствует бинарной номенклатуре, т.е. состоит из двух слов. Например, возбудитель дифтерии пишется как Corynebacterium diphtheriae. Первое слово - название рода и пишется с прописной буквы, второе слово обозначает вид и пишется со строчной буквы.
При повторном упоминании вида родовое название сокращается до начальной буквы, например C. diphtheriae.
Совокупность однородных микроорганизмов, выделенных на питательной среде, характеризующихся сходными морфологическими, тинкториальными (отношение к красителям), культуральными, биохимическими и антигенными свойствами, называется чистой культурой. Чистая культура микроорганизмов, выделенных из определенного источника и отличающихся от других представителей вида, называется штаммом. Близким к понятию «штамм» является понятие «клон». Клон представляет собой совокупность потомков, выращенных из единственной микробной клетки.
Для обозначения некоторых совокупностей микроорганизмов, отличающихся по тем или иным свойствам, употребляется суффикс «вар» (разновидность), поэтому микроорганизмы в зависимости от характера различий обозначают как морфовары (отличие по морфологии), резистентовары (отличие по устойчивости, например, к антибиотикам), серовары (отличие по антигенам), фаговары (отличие по чувствительности к бактериофагам), биовары (отличие по биологическим свойствам), хемовары (отличие по биохимическим свойствам) и т.д.
Раньше основу классификации бактерий составляла особенность строения клеточной стенки. Подразделение бактерий по особенностям строения клеточной стенки связано с возможной вариабельностью их окраски в тот или иной цвет по методу Грама. Согласно этому методу, предложенному в 1884 г. датским ученым Х. Грамом, в зависимости от результатов окраски бактерии делятся на грамположительные, окрашиваемые в сине-фиолетовый цвет, и грамотрицательные, окрашиваемые в красный цвет.
В настоящее время основу классификации составляет степень генетического родства, основанная на изучении строения генома рибосомных РНК (рРНК) (см. главу 5), определении процентного содержания в геноме гуанинцитозиновых пар (ГЦ-пары), построении рестрикционной карты генома, изучении степени гибридизации. Также учитываются и фенотипические показатели: отношение к окраске по Граму, морфологические, культуральные и биохимические свойства, антигенная структура.
Домен Bacteria включает 23 типа, из которых медицинское значение имеют нижеизложенные.
Большинство грамотрицательных бактерий объединены в тип Proteobacteria (по имени греческого бога Proteus, способного принимать различные облики). Тип Proteobacteria подразделен на 5 классов:
Класс Alphaproteobacteria (роды Rickettsia, Orientia, Erlichia, Bartonella, Brucella);
класс Betaproteobacteria (роды Вordetellа, Burholderia, Neisseria, Spirillum);
Класс Gammaproteobacteria (представители семейства Enterobacteriaceae, роды Francisella, Legionella, Coxiella, Pseudomonas, Vibrio);
Класс Deltaproteobacteria (род Bilophila);
Класс Epsilonproteobacteria (роды Campilobacter, Helicobacter). Грамотрицательные бактерии входят также в следующие типы:
тип Chlamydiae (роды Chlamydia, Chlamydophila), тип Spirochaetes (роды Spirocheta, Borrelia, Treponema, Leptospira); тип Bacteroides (роды Bacteroides, Prevotella, Porphyromonas).
Грамположительные бактерии входят в следующие типы:
Тип Firmicutes включает класс Clostridium (роды Clostridium, Peptococcus), класс Bacilli (Listeria, Staphylococcus, Lactobacillus, Streptococcus) и класс Mollicutes (роды Mycoplasma, Ureaplasma), которые являются бактериями, не имеющими клеточную стенку;
тип Actinobacteria (роды Actinomyces, Micrococcus, Corynebacterium, Mycobacterium, Gardnerella, Bifidobacterium, Propionibacterium, Mobiluncus).
2.2.1. Морфологические формы бактерий
Различают несколько основных форм бактерий: кокковидные, палочковидные, извитые и ветвящиеся (рис. 2.1).
Сферические формы, или кокки - шаровидные бактерии размером 0,5-1 мкм, которые по взаимному расположению делятся на микрококки, диплококки, стрептококки, тетракокки, сарцины и стафилококки.
Микрококки (от греч. micros - малый) - отдельно расположенные клетки.
Диплококки (от греч. diploos - двойной), или парные кокки, располагаются парами (пневмококк, гонококк, менингококк), так как клетки после деления не расходятся. Пневмококк (возбудитель пневмонии) имеет с противоположных сторон ланцетовидную форму, а гонококк (возбудитель гонореи) и менингококк (возбу-
 Рис. 2.1.
Формы бактерий
Рис. 2.1.
Формы бактерий
дитель эпидемического менингита) имеют форму кофейных зерен, обращенных вогнутой поверхностью друг к другу.
Стрептококки (от греч. streptos - цепочка) - клетки округлой или вытянутой формы, составляющие цепочку вследствие деления клеток в одной плоскости и сохранения связи между ними в месте деления.
Сарцины (от лат. sarcina - связка, тюк) располагаются в виде пакетов из 8 кокков и более, так как они образуются при делении клетки в трех взаимно перпендикулярных плоскостях.
Стафилококки (от греч. staphyle - виноградная гроздь) - кокки, расположенные в виде грозди винограда в результате деления в разных плоскостях.
Палочковидные бактерии различаются по размерам, форме концов клетки и взаимному расположению клеток. Длина клеток 1-10 мкм, толщина 0,5-2 мкм. Палочки могут быть правильной
(кишечная палочка и др.) и неправильной булавовидной (коринебактерии и др.) формы. К наиболее мелким палочковидным бактериям относятся риккетсии.
Концы палочек могут быть как бы обрезанными (сибиреязвенная бацилла), закругленными (кишечная палочка), заостренными (фузобактерии) или в виде утолщения. В последнем случае палочка похожа на булаву (коринебактерии дифтерии).
Слегка изогнутые палочки называются вибрионами (холерный вибрион). Большинство палочковидных бактерий располагается беспорядочно, так как после деления клетки расходятся. Если после деления клетки остаются связанными общими фрагментами клеточной стенки и не расходятся, то они располагаются под углом друг к другу (коринебактерии дифтерии) или образуют цепочку (сибиреязвенная бацилла).
Извитые формы - спиралевидные бактерии, которые бывают двух видов: спириллы и спирохеты. Спириллы имеют вид штопорообразно извитых клеток с крупными завитками. К патогенным спириллам относятся возбудитель содоку (болезнь укуса крыс), а также кампилобактерии и хеликобактерии, имеющие изгибы, напоминающие крылья летящей чайки. Спирохеты представляют тонкие длинные извитые бактерии, отличающиеся от спирилл более мелкими завитками и характером движения. Особенность их строения описана ниже.
Ветвящиеся - палочковидные бактерии, которые могут иметь разветвление в форме латинской буквы Y, встречающиеся у бифидобактерий, также быть представленными в виде нитевидных разветвленных клеток, способных переплетаться, образуя мицелий, что наблюдается у актиномицет.
2.2.2. Структура бактериальной клетки
Структура бактерий хорошо изучена с помощью электронной микроскопии целых клеток и их ультратонких срезов, а также других методов. Бактериальную клетку окружает оболочка, состоящая из клеточной стенки и цитоплазматической мембраны. Под оболочкой находится протоплазма, состоящая из цитоплазмы с включениями и наследственного аппарата - аналога ядра, называемого нуклеоидом (рис. 2.2). Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили. Некоторые бактерии в неблагоприятных условиях способны образовывать споры.
 Рис. 2.2.
Структура
бактериальной клетки: 1 - капсула; 2 - клеточная стенка; 3 -
цитоплазматическая мембрана; 4 - мезосомы; 5 - нуклеоид; 6 - плазмида; 7
- рибосомы; 8 - включения; 9 - жгутик; 10 - пили (ворсинки)
Рис. 2.2.
Структура
бактериальной клетки: 1 - капсула; 2 - клеточная стенка; 3 -
цитоплазматическая мембрана; 4 - мезосомы; 5 - нуклеоид; 6 - плазмида; 7
- рибосомы; 8 - включения; 9 - жгутик; 10 - пили (ворсинки)
Клеточная стенка - прочная, упругая структура, придающая бактерии определенную форму и вместе с подлежащей цитоплазматической мембраной сдерживающая высокое осмотическое давление в бактериальной клетке. Она участвует в процессе деления клетки и транспорте метаболитов, имеет рецепторы для бактериофагов, бактериоцинов и различных веществ. Наиболее толстая клеточная стенка у грамположительных бактерий (рис. 2.3). Так, если толщина клеточной стенки грамотрицательных бактерий около 15-20 нм, то у грамположительных она может достигать 50 нм и более.
Основу клеточной стенки бактерий составляет пептидогликан. Пептидогликан является полимером. Он представлен параллельными полисахаридными гликановыми цепями, состоящими из повторяющихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных гликозидной связью. Эту связь разрывает лизоцим, являющийся ацетилмурамидазой.
К N-ацетилмурамовой кислоте ковалентными связями присоединен тетрапептид. Тетрапептид состоит из L-аланина, который связан с N-ацетилмурамовой кислотой; D-глутамина, который у грамположительных бактерий соединен с L-лизином, а у грамотри-
 Рис. 2.3.
Схема архитектоники клеточной стенки бактерий
Рис. 2.3.
Схема архитектоники клеточной стенки бактерий
цательных бактерий - с диаминопимелиновой кислотой (ДАП), которая представляет собой предшественник лизина в процессе бактериального биосинтеза аминокислот и является уникальным соединением, присутствующим только у бактерий; 4-й аминокислотой является D-аланин (рис. 2.4).
В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов и белков. Основным компонентом клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90% массы клеточной стенки. Тетрапептиды разных слоев пептидогликана у грамположительных бактерий соединены друг с другом полипептидными цепочками из 5 остатков глицина (пентаглицина), что придает пептидогликану жесткую геометрическую структуру (рис. 2.4, б). С пептидогликаном ктеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. tekhos - стенка), молекулы которых представляют собой цепи из 8-50 остатков глицерола и рибитола, соединенных фосфатными мостиками. Форму и прочность бактериям придает жесткая волокнистая структура многослойного, с поперечными пептидными сшивками пептидогликана.
 Рис. 2.4.
Структура пептидогликана: а - грамотрицательные бактерии; б - грамположительные бактерии
Рис. 2.4.
Структура пептидогликана: а - грамотрицательные бактерии; б - грамположительные бактерии
Способность грамположительных бактерий при окраске по Граму удерживать генциановый фиолетовый в комплексе с йодом (сине-фиолетовая окраска бактерий) связана со свойством многослойного пептидогликана взаимодействовать с красителем. Кроме этого последующая обработка мазка бактерий спиртом вызывает сужение пор в пептидогликане и тем самым задерживает краситель в клеточной стенке.
Грамотрицательные бактерии после воздействия спиртом утрачивают краситель, что обусловлено меньшим количеством пептидогликана (5-10% массы клеточной стенки); они обесцвечиваются спиртом, и при обработке фуксином или сафранином приобретают красный цвет. Это связано с особенностями строения клеточной стенки. Пептидогликан в клеточной стенке грамотрицательных бактерий представлен 1-2 слоями. Тетрапептиды слоев соединены между собой прямой пептидной связью между аминогруппой ДАП одного тетрапептида и карбоксильной группой D-аланина тетрапептида другого слоя (рис. 2.4, а). Кнаружи от пептидогликана расположен слой липопротеина, соединенный с пептидогликаном через ДАП. За ним следует наружная мембрана клеточной стенки.
Наружная мембрана является мозаичной структурой, представленной липополисахаридами (ЛПС), фосфолипидами и белками. Внутренний слой ее представлен фосфолипидами, а в наружном слое расположен ЛПС (рис. 2.5). Таким образом, наружная мем-
 Рис. 2.5.
Структура липополисахарида
Рис. 2.5.
Структура липополисахарида
брана асимметрична. ЛПС наружной мембраны состоит из трех фрагментов:
Липида А - консервативной структуры, практически одинаковой у грамотрицательных бактерий. Липид А состоит из фосфорилированных глюкозоаминовых дисахаридных единиц, к которым прикреплены длинные цепочки жирных кислот (см. рис. 2.5);
Ядра, или стержневой, коровой части (от лат. core - ядро), относительно консервативной олигосахаридной структуры;
Высоковариабельной О-специфической цепи полисахарида, образованной повторяющимися идентичными олигосахаридными последовательностями.
ЛПС заякорен в наружной мембране липидом А, обусловливающим токсичность ЛПС и отождествляемым поэтому с эндотоксином. Разрушение бактерий антибиотиками приводит к освобождению большого количества эндотоксина, что может вызвать у больного эндотоксический шок. От липида А отходит ядро, или стержневая часть ЛПС. Наиболее постоянной частью ядра ЛПС является кетодезоксиоктоновая кислота. О-специфическая полисахаридная цепь, отходящая от стержневой части молекулы ЛПС,
состоящая из повторяющихся олигосахаридных единиц, обусловливает серогруппу, серовар (разновидность бактерий, выявляемая с помощью иммунной сыворотки) определенного штамма бактерий. Таким образом, с понятием ЛПС связаны представления об О-антигене, по которому можно дифференцировать бактерии. Генетические изменения могут привести к дефектам, укорочению ЛПС бактерий и появлению в результате этого шероховатых колоний R-форм, теряющих О-антигенную специфичность.
Не все грамотрицательные бактерии имеют полноценную О-специфическую полисахаридную цепь, состоящую из повторяющихся олигосахаридных единиц. В частности, бактерии рода Neisseria имеют короткий гликолипид, который называется липоолигосахаридом (ЛОС). Он сравним с R-формой, потерявшей О-антигенную специфичность, наблюдаемой у мутантных шероховатых штаммов E. coli. Структура ЛОС напоминает структуру гликосфинголипида цитоплазматической мембраны человека, поэтому ЛОС мимикрирует микроб, позволяя ему избегать иммунного ответа хозяина.
Белки матрикса наружной мембраны пронизывают ее таким образом, что молекулы белка, называемые поринами, окаймляют гидрофильные поры, через которые проходят вода и мелкие гидрофильные молекулы с относительной массой до 700 Д.
Между наружной и цитоплазматической мембраной находится периплазматическое пространство, или периплазма, содержащая ферменты (протеазы, липазы, фосфатазы, нуклеазы, β-лактамазы), а также компоненты транспортных систем.
При нарушении синтеза клеточной стенки бактерий под влиянием лизоцима, пенициллина, защитных факторов организма и других соединений образуются клетки с измененной (часто шаровидной) формой: протопласты - бактерии, полностью лишенные клеточной стенки; сферопласты - бактерии с частично сохранившейся клеточной стенкой. После удаления ингибитора клеточной стенки такие измененные бактерии могут реверсировать, т.е. приобретать полноценную клеточную стенку и восстанавливать исходную форму.
Бактерии сфероили протопластного типа, утратившие способность к синтезу пептидогликана под влиянием антибиотиков или других факторов и способные размножаться, называются L-формами (от названия Института им. Д. Листера, где они впер-
вые были изучены). L-формы могут возникать и в результате мутаций. Они представляют собой осмотически чувствительные, шаровидные, колбовидные клетки различной величины, в том числе и проходящие через бактериальные фильтры. Некоторые L-формы (нестабильные) при удалении фактора, приведшего к изменениям бактерий, могут реверсировать, возвращаясь в исходную бактериальную клетку. L-формы могут образовывать многие возбудители инфекционных болезней.
Цитоплазматическая мембрана при электронной микроскопии ультратонких срезов представляет собой трехслойную мембрану (2 темных слоя толщиной по 2,5 нм каждый разделены светлым - промежуточным). По структуре она похожа на плазмолемму клеток животных и состоит из двойного слоя липидов, главным образом фосфолипидов, с внедренными поверхностными, а также интегральными белками, как бы пронизывающими насквозь структуру мембраны. Некоторые из них являются пермеазами, участвующими в транспорте веществ. В отличие от эукариотических клеток, в цитоплазматической мембране бактериальной клетки отсутствуют стеролы (за исключением микоплазм).
Цитоплазматическая мембрана является динамической структурой с подвижными компонентами, поэтому ее представляют как мобильную текучую структуру. Она окружает наружную часть цитоплазмы бактерий и участвует в регуляции осмотического давления, транспорте веществ и энергетическом метаболизме клетки (за счет ферментов цепи переноса электронов, аденозинтрифосфатазы - АТФазы и др.). При избыточном росте (по сравнению с ростом клеточной стенки) цитоплазматическая мембрана образует инвагинаты - впячивания в виде сложно закрученных мембранных структур, называемые мезосомами. Менее сложно закрученные структуры называются внутрицитоплазматическими мембранами. Роль мезосом и внутрицитоплазматических мембран до конца не выяснена. Предполагают даже, что они являются артефактом, возникающим после приготовления (фиксации) препарата для электронной микроскопии. Тем не менее считают, что производные цитоплазматической мембраны участвуют в делении клетки, обеспечивая энергией синтез клеточной стенки, принимают участие в секреции веществ, спорообразовании, т.е. в процессах с высокой затратой энергии. Цитоплазма занимает основной объем бактери-
альной клетки и состоит из растворимых белков, рибонуклеиновых кислот, включений и многочисленных мелких гранул - рибосом, ответственных за синтез (трансляцию) белков.
Рибосомы бактерий имеют размер около 20 нм и коэффициент седиментации 70S, в отличие от 80S-рибосом, характерных для эукариотических клеток. Поэтому некоторые антибиотики, связываясь с рибосомами бактерий, подавляют синтез бактериального белка, не влияя на синтез белка эукариотических клеток. Рибосомы бактерий могут диссоциировать на две субъединицы: 50S и 30S. рРНК - консервативные элементы бактерий («молекулярные часы» эволюции). 16S-рРНК входит в состав малой субъединицы рибосом, а 23S-рРНК - в состав большой субъединицы рибосом. Изучение 16S рРНК является основой геносистематики, позволяя оценить степень родства организмов.
В цитоплазме имеются различные включения в виде гранул гликогена, полисахаридов, β-оксимасляной кислоты и полифосфатов (волютин). Они накапливаются при избытке питательных веществ в окружающей среде и выполняют роль запасных веществ для питания и энергетических потребностей.
Волютин обладает сродством к основным красителям и легко выявляется с помощью специальных методов окраски (например, по Нейссеру) в виде метахроматических гранул. Толуидиновым синим или метиленовым голубым волютин окрашивается в краснофиолетовый цвет, а цитоплазма бактерии - в синий. Характерное расположение гранул волютина выявляется у дифтерийной палочки в виде интенсивно прокрашивающихся полюсов клетки. Метахроматическое окрашивание волютина связано с высоким содержанием полимеризованного неорганического полифосфата. При электронной микроскопии они имеют вид электронноплотных гранул размером 0,1-1 мкм.
Нуклеоид - эквивалент ядра у бактерий. Он расположен в центральной зоне бактерий в виде двунитевой ДНК, плотно уложенной наподобие клубка. Нуклеоид бактерий, в отличие от эукариот, не имеет ядерной оболочки, ядрышка и основных белков (гистонов). У большинства бактерий содержится одна хромосома, представленная замкнутой в кольцо молекулой ДНК. Но у некоторых бактерий имеются две хромосомы кольцевой формы (V. cholerae) и линейные хромосомы (см. раздел 5.1.1). Нуклеоид выявляется в световом микроскопе после окраски специфическими для ДНК
методами: по Фельгену или по Романовскому-Гимзе. На электронограммах ультратонких срезов бактерий нуклеоид имеет вид светлых зон с фибриллярными, нитевидными структурами ДНК, связанной определенными участками с цитоплазматической мембраной или мезосомой, участвующими в репликации хромосомы.
Кроме нуклеоида, в бактериальной клетке имеются внехромосомные факторы наследственности - плазмиды (см. раздел 5.1.2), представляющие собой ковалентно замкнутые кольца ДНК.
Капсула, микрокапсула, слизь. Капсула - слизистая структура толщиной более 0,2 мкм, прочно связанная с клеточной стенкой бактерий и имеющая четко очерченные внешние границы. Капсула различима в мазках-отпечатках из патологического материала. В чистых культурах бактерий капсула образуется реже. Она выявляется при специальных методах окраски мазка по Бурри- Гинсу, создающих негативное контрастирование веществ капсулы: тушь создает темный фон вокруг капсулы. Капсула состоит из полисахаридов (экзополисахаридов), иногда из полипептидов, например у сибиреязвенной бациллы она состоит из полимеров D-глутаминовой кислоты. Капсула гидрофильна, включает большое количество воды. Она препятствует фагоцитозу бактерий. Капсула антигенна: антитела к капсуле вызывают ее увеличение (реакция набухания капсулы).
Многие бактерии образуют микрокапсулу - слизистое образование толщиной менее 0,2 мкм, выявляемое лишь при электронной микроскопии.
От капсулы следует отличать слизь - мукоидные экзополисахариды, не имеющие четких внешних границ. Слизь растворима в воде.
Мукоидные экзополисахариды характерны для мукоидных штаммов синегнойной палочки, часто встречающихся в мокроте больных кистозным фиброзом. Бактериальные экзополисахариды участвуют в адгезии (прилипании к субстратам); их еще называют гликокаликсом.
Капсула и слизь предохраняют бактерии от повреждений, высыхания, так как, являясь гидрофильными, хорошо связывают воду, препятствуют действию защитных факторов макроорганизма и бактериофагов.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие на-
чало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из трех частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (одна пара дисков у грамположительных и две пары у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем - ротором, вращающим жгутик. В качестве источника энергии используется разность протонных потенциалов на цитоплазматической мембране. Механизм вращения обеспечивает протонная АТФ-синтетаза. Скорость вращения жгутика может достигать 100 об/с. При наличии у бактерии нескольких жгутиков они начинают синхронно вращаться, сплетаясь в единый пучок, образующий своеобразный пропеллер.
Жгутики состоят из белка - флагеллина (flagellum - жгутик), являющегося антигеном - так называемый Н-антиген. Субъединицы флагеллина закручены в виде спирали.
Число жгутиков у бактерий разных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен, отходящих по периметру бактерии (перитрих), у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Жгутики выявляют с помощью электронной микроскопии препаратов, напыленных тяжелыми металлами, или в световом микроскопе после обработки специальными методами, основанными на протравливании и адсорбции различных веществ, приводящих к увеличению толщины жгутиков (например, после серебрения).
Ворсинки, или пили (фимбрии) - нитевидные образования, более тонкие и короткие (3-10 нм * 0,3-10 мкм), чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина. Известно несколько типов пилей. Пили общего типа отвечают за прикрепления к субстрату, питание и водно-солевой обмен. Они многочисленны - несколько сотен на клетку. Половые пили (1-3 на клетку) создают контакт между клетками, осуществляя между ними передачу генетической информации путем конъюгации (см. главу 5). Особый интерес представляют пили IV типа, у которых концы обладают гидрофобностью, в результате чего они закручиваются, эти пили называют еще кудряшками. Располага-
ются они по полюсам клетки. Эти пили встречаются у патогенных бактерий. Они обладают антигенными свойствами, осуществляют контакт бактерии с клеткой-хозяином, участвуют в образовании биопленки (см. главу 3). Многие пили являются рецепторами для бактериофагов.
Споры - своеобразная форма покоящихся бактерий с грамположительным типом строения клеточной стенки. Спорообразующие бактерии рода Bacillus, у которых размер споры не превышает диаметр клетки, называются бациллами. Спорообразующие бактерии, у которых размер споры превышает диаметр клетки, отчего они принимают форму веретена, называются клостридиями, например бактерии рода Clostridium (от лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нельсена в красный, а вегетативная клетка - в синий цвет.
Спорообразование, форма и расположение спор в клетке (вегетативной) являются видовым свойством бактерий, что позволяет отличать их друг от друга. Форма спор бывает овальной и шаровидной, расположение в клетке - терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулизма, газовой гангрены) и центральное (у сибиреязвенной бациллы).
Процесс спорообразования (споруляция) проходит ряд стадий, в течение которых часть цитоплазмы и хромосома бактериальной вегетативной клетки отделяются, окружаясь врастающей цитоплазматической мембраной, - образуется проспора.
В протопласте проспоры находятся нуклеоид, белоксинтезирующая система и система получения энергии, основанная на гликолизе. Цитохромы отсутствуют даже у аэробов. Не содержится АТФ, энергия для прорастания сохраняется в форме 3-глицеринфосфата.
Проспору окружают две цитоплазматические мембраны. Слой, окружающий внутреннюю мембрану споры, называется стенкой споры, он состоит из пептидогликана и является главным источником клеточной стенки при прорастании споры.
Между наружной мембраной и стенкой споры формируется толстый слой, состоящий из пептидогликана, имеющего много сшивок, - кортекс.
Кнаружи от внешней цитоплазматической мембраны расположена оболочка споры, состоящая из кератиноподобных белков, со-
держащих множественные внутримолекулярные дисульфидные связи. Эта оболочка обеспечивает резистентность к химическим агентам. Споры некоторых бактерий имеют дополнительный покров - экзоспориум липопротеиновой природы. Таким образом формируется многослойная плохо проницаемая оболочка.
Спорообразование сопровождается интенсивным потреблением проспорой, а затем и формирующейся оболочкой споры дипиколиновой кислоты и ионов кальция. Спора приобретает термоустойчивость, которую связывают с наличием в ней дипиколината кальция.
Спора долго может сохраняться из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизма. В почве, например, возбудители сибирской язвы и столбняка могут сохраняться десятки лет.
В благоприятных условиях споры прорастают, проходя три последовательные стадии: активации, инициации, вырастания. При этом из одной споры образуется одна бактерия. Активация - это готовность к прорастанию. При температуре 60-80 °С спора активируется для прорастания. Инициация прорастания длится несколько минут. Стадия вырастания характеризуется быстрым ростом, сопровождающимся разрушением оболочки и выходом проростка.
2.2.3. Особенности строения спирохет, риккетсий, хламидий, актиномицет и микоплазм
Спирохеты - тонкие длинные извитые бактерии. Они состоят из наружной мембранной клеточной стенки, которая окружает цитоплазматический цилиндр. Поверх наружной мембраны располагается прозрачный чехол гликозаминогликановой природы. Под наружной мембранной клеточной стенки располагаются фибриллы, закручивающиеся вокруг цитоплазматического цилиндра, придавая бактериям винтообразную форму. Фибриллы прикреплены к концам клетки и направлены навстречу друг другу. Число и расположение фибрилл варьируют у разных видов. Фибриллы участвуют в передвижении спирохет, придавая клеткам вращательное, сгибательное и поступательное движение. При этом спирохеты образуют петли, завитки, изгибы, которые названы вторичными завитками. Спирохеты плохо воспринимают красители. Обычно их окрашивают по Романовскому-Гимзе или серебрением. В живом
виде спирохеты исследуют с помощью фазово-контрастной или темнопольной микроскопии.
Спирохеты представлены тремя родами, патогенными для человека: Treponema, Borrelia, Leptospira.
Трепонемы (род Treponema) имеют вид тонких штопорообразно закрученных нитей с 8-12 равномерными мелкими завитками. Вокруг протопласта трепонем расположены 3-4 фибриллы (жгутики). В цитоплазме имеются цитоплазматические филаменты. Патогенными представителями являются Т. pallidum - возбудитель сифилиса, T. pertenue - возбудитель тропической болезни - фрамбезии. Имеются и сапрофиты - обитатели полости рта человека, ила водоемов.
Боррелии (род Borrelia), в отличие от трепонем, более длинные, имеют по 3-8 крупных завитков и 7-20 фибрилл. К ним относятся возбудитель возвратного тифа (В. recurrentis) и возбудители болезни Лайма (В. burgdorferi) и других заболеваний.
Лептоспиры (род Leptospira) имеют завитки неглубокие и частые в виде закрученной веревки. Концы этих спирохет изогнуты наподобие крючков с утолщениями на концах. Образуя вторичные завитки, они приобретают вид букв S или С; имеют две осевые фибриллы. Патогенный представитель L. interrogans вызывает лептоспироз при попадании в организм с водой или пищей, приводя к кровоизлияниям и желтухе.
Риккетсии обладают независимым от клетки хозяина метаболизмом, однако, возможно, они получают от клетки хозяина макроэргические соединения для своего размножения. В мазках и тканях их окрашивают по Романовскому-Гимзе, по Маккиавелло- Здродовскому (риккетсии красного цвета, а инфицированные клетки - синего).
У человека риккетсии вызывают эпидемический сыпной тиф (R. prowazekii), клещевой риккетсиоз (R. sibirica), пятнистую лихорадку Скалистых гор (R. rickettsii) и другие риккетсиозы.
Строение их клеточной стенки напоминает таковую грамотрицательных бактерий, хотя имеются отличия. Она не содержит типичного пептидогликана: в его составе полностью отсутствует N-ацетилмурамовая кислота. В состав клеточной стенки входит двойная наружная мембрана, которая включает липополисахарид и белки. Несмотря на отсутствие пептидогликана, клеточная стенка хламидий обладает ригидностью. Цитоплазма клетки ограничена внутренней цитоплазматической мембраной.
Основным методом выявления хламидий является окраска по Романовскому-Гимзе. Цвет окраски зависит от стадии жизненного цикла: элементарные тельца окашиваются в пурпурный цвет на фоне голубой цитоплазмы клетки, ретикулярные тельца - в голубой цвет.
У человека хламидии вызывают поражения глаз (трахома, конъюнктивит), урогенитального тракта, легких и др.
Актиномицеты - ветвящиеся, нитевидные или палочковидные грамположительные бактерии. Свое название (от греч. actis - луч, mykes - гриб) они получили в связи с образованием в пораженных тканях друз - гранул из плотно переплетенных нитей в виде
лучей, отходящих от центра и заканчивающихся колбовидными утолщениями. Актиномицеты, как и грибы, образуют мицелий - нитевидные переплетающиеся клетки (гифы). Они формируют субстратный мицелий, образующийся в результате врастания клеток в питательную среду, и воздушный, растущий на поверхности среды. Актиномицеты могут делиться путем фрагментации мицелия на клетки, похожие на палочковидные и кокковидные бактерии. На воздушных гифах актиномицетов образуются споры, служащие для размножения. Споры актиномицетов обычно не термостойки.
Общую филогенетическую ветвь с актиномицетами образуют так называемые нокардиоподобные (нокардиоформные) актиномицеты - собирательная группа палочковидных бактерий неправильной формы. Их отдельные представители образуют ветвящиеся формы. К ним относят бактерии родов Corynebacterium, Mycobacterium, Nocardia и др. Нокардиоподобные актиномицеты отличаются наличием в клеточной стенке сахаров арабинозы, галактозы, а также миколовых кислот и больших количеств жирных кислот. Миколовые кислоты и липиды клеточных стенок обусловливают кислотоустойчивость бактерий, в частности микобактерий туберкулеза и лепры (при окраске по Цилю-Нельсену они имеют красный цвет, а некислотоустойчивые бактерии и элементы ткани, мокроты - синий цвет).
Патогенные актиномицеты вызывают актиномикоз, нокардии - нокардиоз, микобактерии - туберкулез и лепру, коринебактерии - дифтерию. Сапрофитные формы актиномицетов и нокардиоподобных актиномицетов широко распространены в почве, многие из них являются продуцентами антибиотиков.
Микоплазмы - мелкие бактерии (0,15-1 мкм), окруженные только цитоплазматической мембраной, содержащей стеролы. Они относятся к классу Mollicutes. Из-за отсутствия клеточной стенки микоплазмы осмотически чувствительны. Имеют разнообразную форму: кокковидную, нитевидную, колбовидную. Эти формы видны при фазово-контрастной микроскопии чистых культур микоплазм. На плотной питательной среде микоплазмы образуют колонии, напоминающие яичницу-глазунью: центральная непрозрачная часть, погруженная в среду, и просвечивающая периферия в виде круга.
Микоплазмы вызывают у человека атипичную пневмонию (Mycoplasma pneumoniae) и поражения мочеполового тракта
(М. hominis и др.). Микоплазмы вызывают заболевания не только у животных, но и у растений. Достаточно широко распространены и непатогенные представители.
2.3. Строение и классификация грибов
Грибы относятся к домену Eukarya, царству Fungi (Mycota, Mycetes). Недавно грибы и простейшие были разделены на самостоятельные царства: царство Eumycota (настоящие грибы), царство Chromista и царство Protozoa. Некоторые микроорганизмы, ранее считавшиеся грибами или простейшими, были перемещены в новое царство Chromista (хромовики). Грибы - многоклеточные или одноклеточные нефотосинтезирующие (бесхлорофильные) эукариотические микроорганизмы с толстой клеточной стенкой. Они имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и многослойную ригидную клеточную стенку, состоящую из нескольких типов полисахаридов (маннаны, глюканы, целлюлоза, хитин), а также белка, липидов и др. Некоторые грибы образуют капсулу. Цитоплазматическая мембрана содержит гликопротеины, фосфолипиды и эргостеролы (в отличие от холестерина - главного стерола тканей млекопитающих). Большинство грибов - облигатные или факультативные аэробы.
Грибы широко распространены в природе, особенно в почве. Некоторые грибы содействуют производству хлеба, сыра, молочнокислых продуктов и алкоголя. Другие грибы продуцируют антимикробные антибиотики (например, пенициллин) и иммунодепрессивные лекарства (например, циклоспорин). Грибы используют генетики и молекулярные биологи для моделирования различных процессов. Фитопатогенные грибы наносят значительный ущерб сельскому хозяйству, вызывая грибковые болезни злаковых растений и зерна. Инфекции, вызываемые грибами, называются микозами. Различают гифальные и дрожжевые грибы.
Гифальные (плесневые) грибы, или гифомицеты, состоят из тонких нитей толщиной 2-50 мкм, называемых гифами, которые сплетаются в грибницу или мицелий (плесень). Тело гриба называется талломом. Различают демациевые (пигментированные - коричневые или черные) и гиалиновые (непигментированные) гифомицеты. Гифы, врастающие в питательный субстрат, отвечают за питание гриба и называются вегетативными гифами. Гифы, ра-
стущие над поверхностью субстрата, называются воздушными или репродуктивными гифами (отвечают за размножение). Колонии из-за воздушного мицелия имеют пушистый вид.
Различают низшие и высшие грибы: гифы высших грибов разделены перегородками, или септами с отверстиями. Гифы низших грибов не имеют перегородок, представляя собой многоядерные клетки, называемые ценоцитными (от греч. koenos - единый, общий).
Дрожжевые грибы (дрожжи) в основном представлены отдельными овальными клетками диаметром 3-15 мкм, а их колонии, в отличие от гифальных грибов, имеют компактный вид. По типу полового размножения они распределены среди высших грибов - аскомицет и базидиомицет. При бесполом размножении дрожжи образуют почки или делятся. Могут образовывать псевдогифы и ложный мицелий (псевдомицелий) в виде цепочек удлиненных клеток - «сарделек». Грибы, аналогичные дрожжам, но не имеющие полового способа размножения, называют дрожжеподобными. Они размножаются только бесполым способом - почкованием или делением. Понятия «дрожжеподобные грибы» часто идентифицируют с понятием «дрожжи».
Многие грибы обладают диморфизмом - способностью к гифальному (мицелиальному) или дрожжеподобному росту в зависимости от условий культивирования. В инфицированном организме они растут в виде дрожжеподобных клеток (дрожжевая фаза), а на питательных средах образуют гифы и мицелий. Диморфизм связан с температурным фактором: при комнатной температуре образуется мицелий, а при 37 °С (при температуре тела человека) - дрожжеподобные клетки.
Грибы размножаются половым или бесполым способом. Половое размножение грибов происходит с образованием гамет, половых спор и других половых форм. Половые формы называются телеоморфами.
Бесполое размножение грибов происходит с образованием соответствующих форм, называемых анаморфами. Такое размножение происходит почкованием, фрагментацией гиф и бесполыми спорами. Эндогенные споры (спорангиоспоры) созревают внутри округлой структуры - спорангия. Экзогенные споры (конидии) формируются на кончиках плодоносящих гиф, так называемых конидиеносцах.
Различают разнообразые конидии. Артроконидии (артроспоры), или таллоконидии, образуются при равномерном септировании и расчленении гиф, а бластоконидии образуются в результате почкования. Небольшие одноклеточные конидии называются микроконидиями, большие многоклеточные конидии - макроконидиями. К бесполым формам грибов относят также хламидоконидии, или хламидоспоры (толстостенные крупные покоящиеся клетки или комплекс мелких клеток).
Различают совершенные и несовершенные грибы. Совершенные грибы имеют половой способ размножения; к ним относят зигомицеты (Zygomycota), аскомицеты (Ascomycota) и базидиомицеты (Basidiomycota). Несовершенные грибы имеют только бесполый способ размножения; к ним относят формальный условный тип/ группу грибов - дейтеромицеты (Deiteromycota).
Зигомицеты относятся к низшим грибам (мицелий несептированный). Они включают представителей родов Mucor, Rhizopus, Rhizomucor, Absidia, Basidiobolus, Conidiobolus. Распространены в почве и воздухе. Могут вызывать зигомикоз (мукоромикоз) легких, головного мозга и других органов человека.
При бесполом размножении зигомицет на плодоносящей гифе (спорангиеносце) образуется спорангий - шаровидное утолщение с оболочкой, содержащее многочисленные спорангиоспоры (рис. 2.6, 2.7). Половое размножение у зигомицетов происходит с помощью зигоспор.
Аскомицеты (сумчатые грибы) имеют септированный мицелий (кроме одноклеточных дрожжей). Свое название они получили от основного органа плодоношения - сумки, или аска, содержащего 4 или 8 гаплоидных половых спор (аскоспор).
К аскомицетам относятся отдельные представители (телеоморфы) родов Aspergillus и Penicillium. Большинство грибов родов Aspergillus, Penicillium являются анаморфами, т.е. размножаются только беспо-
 Рис. 2.6.
Грибы рода Mucor
(рис. А.С. Быкова)
Рис. 2.6.
Грибы рода Mucor
(рис. А.С. Быкова)
 Рис. 2.7.
Грибы рода Rhizopus.
Развитие спорангия, спорангиоспор и ризоидов
Рис. 2.7.
Грибы рода Rhizopus.
Развитие спорангия, спорангиоспор и ризоидов
лым путем с помощью бесполых спор - конидий (рис. 2.8, 2.9) и должны быть отнесены по этому признаку к несовершенным грибам. У грибов рода Aspergillus на концах плодоносящих гиф, конидиеносцах, имеются утолщения - стеригмы, фиалиды, на которых образуются цепочки конидий («леечная плесень»).
У грибов рода Penicillium (кистевик) плодоносящая гифа напоминает кисточку, так как из нее (на конидиеносце) образуются утолщения, разветвляющиеся на более мелкие структуры - стеригмы, фиалиды, на которых находятся цепочки конидий. Некоторые виды аспергилл могут вызывать аспергиллезы и афлатоксикозы, пенициллы могут вызывать пенициллиозы.
Представителями аскомицетов являются телеоморфы родов Trichophyton, Microsporum, Histoplasma, Blastomyces, а также дрож-
 Рис. 2.8.
Грибы рода Penicillium.
Цепочки конидий отходят от фиалид
Рис. 2.8.
Грибы рода Penicillium.
Цепочки конидий отходят от фиалид
 Рис. 2.9.
Грибы рода Aspergillus fumigatus.
От фиалид отходят цепочки конидий
Рис. 2.9.
Грибы рода Aspergillus fumigatus.
От фиалид отходят цепочки конидий
Базидиомицеты включают шляпочные грибы. Они имеют септированный мицелий и образуют половые споры - базидиоспоры путем отшнуровывания от базидия - концевой клетки мицелия, гомологичной аску. К базидиомицетам относятся некоторые дрожжи, например телеоморфы Cryptococcus neoformans.
Дейтеромицеты являются несовершенными грибами (Fungi imperfecti, анаморфные грибы, конидиальные грибы). Это условный, формальный таксон грибов, объединяющий грибы, не имеющие полового размножения. Недавно вместо термина «дейтеромицеты» предложен термин «митоспоровые грибы» - грибы, размножающиеся неполовыми спорами, т.е. путем митоза. При установлении факта полового размножения несовершенных грибов их переносят в один из известных типов - Ascomycota или Basidiomycota, присваивая название телеоморфной формы. Дейтеромицеты имеют септированный мицелий, размножаются только путем бесполого формирования конидий. К дейтеромицетам относятся несовершенные дрожжи (дрожжеподобные грибы), например некоторые грибы рода Candida, поражающие кожу, слизистые оболочки и внутренние органы (кандидоз). Они имеют овальную форму, диаметр 2-5 мкм, делятся почкованием, образуют псевдогифы (псевдомицелий) в виде цепочек из удлиненных клеток, иногда образуют гифы. Для Candida albicans характерно образование хламидоспор (рис. 2.10). К дейтеромицетам относят также другие грибы, не имеющие полового способа размножения, относящиеся к родам Epidermophyton, Coccidioides, Paracoccidioides, Sporothrix, Aspergillus, Phialophora, Fonsecaeа, Exophiala, Cladophialophora, Bipolaris, Exerohilum, Wangiella, Alrernaria и др.
 Рис. 2.10.
Грибы рода Candida albicans
(рис. А.С. Быкова)
Рис. 2.10.
Грибы рода Candida albicans
(рис. А.С. Быкова)
2.4. Строение и классификация простейших
Простейшие относятся к домену Eukarya, царству животных (Animalia), подцарству Protozoa. Недавно предложено выделить простейшие в ранг царства Protozoa.
Клетка простейших окружена мембраной (пелликулой) - аналогом цитоплазматической мембраны клеток животных. Она имеет ядро с ядерной оболочкой и ядрышком, цитоплазму, содержащую эндоплазматический ретикулум, митохондрии, лизосомы и рибосомы. Размеры простейших колеблются от 2 до 100 мкм. При окраске по Романовскому-Гимзе ядро простейших имеет красный, а цитоплазма - голубой цвет. Простейшие передвигаются с помощью жгутиков, ресничек или псевдоподий, некоторые из них имеют пищеварительные и сократительные (выделительные) вакуоли. Они могут питаться в результате фагоцитоза или образования особых структур. По типу питания они разделяются на гетеротрофы и аутотрофы. Многие простейшие (дизентерийная амеба, лямблии, трихомонады, лейшмании, балантидии) могут расти на питательных средах, содержащих нативные белки и аминокислоты. Для их культивирования используют также культуры клеток, куриные эмбрионы и лабораторных животных.
Простейшие размножаются бесполым путем - двойным или множественным (шизогония) делением, а некоторые и половым путем (спорогония). Одни простейшие размножаются внеклеточно (лямблии), а другие - внутриклеточно (плазмодии, токсоплазма, лейшмании). Жизненный цикл простейших характеризуется стадийностью - образованием стадии трофозоита и стадии цисты. Цисты - покоящиеся стадии, устойчивые к изменению температуры и влажности. Кислотоустойчивостью отличаются цисты Sarcocystis, Cryptosporidium и Isospora.
Ранее простейшие, вызывающие заболевания у человека, были представлены 4 типами 1 (Sarcomastigophora, Apicomplexa, Ciliophora, Microspora). Эти типы недавно реклассифицированы на большее количество, появились новые царства - Protozoa и Chromista (табл. 2.2). В новое царство Chromista (хромовики) вошли некоторые простейшие и грибы (бластоцисты, оомицеты и Rhinosporidium seeberi). Царство Protozoa включает амебы, жгутиконосцы, споровики и реснитчатые. Они подразделены на различные типы, среди которых различают амебы, жгутиконосцы, споровики и реснитчатые.
Таблица 2.2. Представители царств Protozoa и Chromista, имеющие медицинское значение
 1 Тип Sarcomastigophora
состоял из подтипов Sarcodina
и Mastigophora.
Подтип Sarcodina
(саркодовые) включал дизентерийную амебу, а подтип Mastigophora
(жгутиконосцы) - трипаносомы, лейшмании, лямблию и трихомонады. Тип Apicomplexa
включал класс Sporozoa
(споровики), куда входили плазмодии малярии, токсоплазма, криптоспоридии и др. Тип Ciliophora
включает балантидии, а тип Microspora
- микроспоридии.
1 Тип Sarcomastigophora
состоял из подтипов Sarcodina
и Mastigophora.
Подтип Sarcodina
(саркодовые) включал дизентерийную амебу, а подтип Mastigophora
(жгутиконосцы) - трипаносомы, лейшмании, лямблию и трихомонады. Тип Apicomplexa
включал класс Sporozoa
(споровики), куда входили плазмодии малярии, токсоплазма, криптоспоридии и др. Тип Ciliophora
включает балантидии, а тип Microspora
- микроспоридии.
Окончание табл. 2.2
 К амебам относятся возбудитель амебиаза человека - амебной дизентерии (Entamoeba histolytica),
свободно
живущие и непатогенные амебы (кишечная амеба и др.). Амебы размножаются
бинарно бесполым путем. Их жизненный цикл состоит из стадии трофозоита
(растущая, подвижная клетка, малоустойчивая) и стадии цисты. Трофозоиты
передвигаются с помощью псевдоподий, которые захватывают и погружают в
цитоплазму питательные вещества. Из
К амебам относятся возбудитель амебиаза человека - амебной дизентерии (Entamoeba histolytica),
свободно
живущие и непатогенные амебы (кишечная амеба и др.). Амебы размножаются
бинарно бесполым путем. Их жизненный цикл состоит из стадии трофозоита
(растущая, подвижная клетка, малоустойчивая) и стадии цисты. Трофозоиты
передвигаются с помощью псевдоподий, которые захватывают и погружают в
цитоплазму питательные вещества. Из
трофозоита образуется циста, устойчивая к внешним факторам. Попав в кишечник, она превращается в трофозоит.
Жгутиконосцы характеризуются наличием жгутиков: у лейшманий один жгутик, у трихомонад 4 свободных жгутика и один жгутик, соединенный с короткой ундулирующей мембраной. Ими являются:
Жгутиконосцы крови и тканей (лейшмании - возбудители лейшманиозов; трипаносомы - возбудители сонной болезни и болезни Шагаса);
Жгутиконосцы кишечника (лямблия - возбудитель лямблиоза);
Жгутиконосцы мочеполового тракта (трихомонада влагалищная - возбудитель трихомоноза).
Реснитчатые представлены балантидиями, которые поражают толстую кишку человека (балантидиазная дизентерия). Балантидии имеют стадию трофозоита и цисты. Трофозоит подвижен, имеет многочисленные реснички, более тонкие и короткие, чем жгутики.
2.5. Строение и классификация вирусов
Вирусы - мельчайшие микробы, относящиеся к царству Virae (от лат. virus - яд). Они не имеют клеточного строения и состоят
Структуру вирусов из-за их малых размеров изучают с помощью электронной микроскопии как вирионов, так и их ультратонких срезов. Размеры вирусов (вирионов) определяют напрямую с помощью электронной микроскопии или косвенно методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Размер вирусов колеблется от 15 до 400 нм (1 нм равен 1/1000 мкм): к маленьким вирусам, размер которых сходен с размером рибосом, относят парвовирусы и вирус полиомиелита, а к наиболее крупным - вирус натуральной оспы (350 нм). Вирусы отличаются по форме вирионов, которые имеют вид палочек (вирус табачной мозаики), пули (вирус бешенства), сферы (вирусы полиомиелита, ВИЧ), нити (филовирусы), сперматозоида (многие бактериофаги).
Вирусы поражают воображение своим разнообразием структуры и свойств. В отличие от клеточных геномов, которые содержат однородную двунитевую ДНК, вирусные геномы чрезвычайно разнообразны. Различают ДНК- и РНК-содержащие вирусы, которые гаплоидны, т.е. имеют один набор генов. Диплоидный геном имеют только ретровирусы. Геном вирусов содержит от 6 до 200 генов и представлен различными видами нуклеиновых кислот: двунитевыми, однонитевыми, линейными, кольцевыми, фрагментированными.
Среди однонитевых РНК-содержащих вирусов различают геномные плюс-нить РНК и минус-нить РНК (полярность РНК). Плюс-нить (позитивная нить) РНК этих вирусов, кроме геномной (наследственной) функции, выполняет функцию информационной, или матричной РНК (иРНК, или мРНК); она является матрицей для белкового синтеза на рибосомах инфицированной клетки. Плюс-нить РНК является инфекционной: при введении в чувствительные клетки она способна вызвать инфекционный про-
цесс. Минус-нить (негативная нить) РНК-содержащих вирусов выполняет только наследственную функцию; для синтеза белка на минус-нити РНК синтезируется комплементарная ей нить. У некоторых вирусов РНК-геном является амбиполярным (ambisense от греч. амби - с обеих сторон, двойная комплементарность), т.е. содержит плюс- и минус-сегменты РНК.
Различают простые вирусы (например, вирус гепатита А) и сложные вирусы (например, вирусы гриппа, герпеса, коронавирусы).
Простые, или безоболочечные, вирусы имеют только нуклеиновую кислоту, связанную с белковой структурой, называемой капсидом (от лат. capsa - футляр). Протеины, связанные с нуклеиновой кислотой, известны как нуклеопротеины, а ассоциация вирусных протеинов капсида вируса с вирусной нуклеиновой кислотой названа нуклеокапсидом. Некоторые простые вирусы могут формировать кристаллы (например, вирус ящура).
Капсид включает повторяющиеся морфологические субъединицы - капсомеры, скомпонованные из нескольких полипептидов. Нуклеиновая кислота вириона, связываясь с капсидом, образует нуклеокапсид. Капсид защищает нуклеиновую кислоту от деградации. У простых вирусов капсид участвует в прикреплении (адсорбции) к клетке хозяина. Простые вирусы выходят из клетки в результате ее разрушения (лизиса).
Сложные, или оболочечные, вирусы (рис. 2.11), кроме капсида, имеют мембранную двойную липопротеиновую оболочку (синоним: суперкапсид, или пеплос), которая приобретается путем почкования вириона через мембрану клетки, например через плазматическую мембрану, мембрану ядра или мембрану эндоплазматического ретикулума. На оболочке вируса расположены гликопротеиновые шипы,
или шипики, пепломеры. Разрушение оболочки эфиром и другими растворителями инактивирует сложные вирусы. Под оболочкой некоторых вирусов находится матриксный белок (М-белок).
Вирионы имеют спиральный, икосаэдрический (кубический) или сложный тип симметрии капсида (нуклеокапсида). Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида (например, у вирусов гриппа, коронавирусов): капсомеры уложены по спирали вместе с нуклеиновой кислотой. Икосаэдрический тип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вируса герпеса).
Капсид и оболочка (суперкапсид) защищают вирионы от воздействия окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) своими рецепторными белками с опреде-
 Рис. 2.11.
Строение оболочечных вирусов с икосаэдрическим (а) и спиральным (б) капсидом
Рис. 2.11.
Строение оболочечных вирусов с икосаэдрическим (а) и спиральным (б) капсидом
ленными клетками, а также антигенные и иммуногенные свойства вирионов.
Внутренние структуры вирусов называют сердцевиной. У аденовирусов сердцевина состоит из гистоноподобных белков, связанных с ДНК, у реовирусов - из белков внутреннего капсида.
Лауреат Нобелевской премии Д. Балтимор предложил систему балтиморской классификации, основанной на механизме синтеза мРНК. Эта классификация размещает вирусы в 7 группах (табл. 2.3). Международный комитет на таксономии вирусов (ICTV) принял универсальную систему классификации, которая использует такие таксономические категории, как семейство (название оканчивается на viridae), подсемейство (название оканчивается на virinae), род (название оканчивается на virus). Вид вируса не получил биноминального названия, как у бактерий. Вирусы классифицируют по типу нуклеиновой кислоты (ДНК или РНК), ее структуре и количеству нитей. Они имеют двунитевые или однонитевые нуклеиновые кислоты; позитивную (+), негативную (-) полярность нуклеиновой кислоты или смешанную полярность нуклеиновой кислоты, амбиполярную (+, -); линейную или циркулярную нуклеиновую кислоту; фрагментированную или нефрагментированную нуклеиновую кислоту. Учитывают также размер и морфологию вирионов, количество капсомеров и тип симметрии нуклеокапсида, наличие оболочки (суперкапсида), чувствительность к эфиру и дезоксихолату, место размножения в клетке, антигенные свойства и др.
Таблица 2.3. Основные вирусы, имеющие медицинское значение
 Продолжение табл. 2.3
Продолжение табл. 2.3
 Окончание табл. 2.3
Окончание табл. 2.3
 Вирусы
поражают животных, бактерии, грибы и растения. Являясь основными
возбудителями инфекционных заболеваний человека, вирусы также участвуют в
процессах канцерогенеза, могут передаваться различными путями, в том
числе через плаценту (вирус краснухи, цитомегаловирус и др.), поражая
плод человека. Они могут приводить и к постинфекционным осложнениям -
развитию миокардитов, панкреатитов, иммунодефицитов и др.
Вирусы
поражают животных, бактерии, грибы и растения. Являясь основными
возбудителями инфекционных заболеваний человека, вирусы также участвуют в
процессах канцерогенеза, могут передаваться различными путями, в том
числе через плаценту (вирус краснухи, цитомегаловирус и др.), поражая
плод человека. Они могут приводить и к постинфекционным осложнениям -
развитию миокардитов, панкреатитов, иммунодефицитов и др.
К неклеточным формам жизни, кроме вирусов, относят прионы и вироиды. Вироиды - небольшие молекулы кольцевой, суперспирализованной РНК, не содержащие белок и вызывающие заболевания растений. Патологические прионы - инфекционные белковые частицы, вызывающие особые конформационные болезни в результате изменения структуры нормального клеточного прионового протеина (PrP c ), который имеется в организме животных и человека. PrP с выполняет регуляторные функции. Его кодирует нормальный прионовый ген (PrP-ген), расположенный в коротком плече 20-й хромосомы человека. Прионные болезни протекают по типу трансмиссивных губкообразных энцефалопатий (болезнь Крейтцфельда-Якоба, куру и др.). При этом прионный протеин приобретает другую, инфекционную форму, обозначаемую как PrP sc (sc от scrapie - скрепи - прионная инфекция овец и коз). Этот инфекционный прионный протеин имеет вид фибрилл и отличается от нормального прионного протеина третичной или четвертичной структурой.
Задания для самоподготовки (самоконтроля)
А. Отметьте микробы, являющиеся прокариотами:
2. Вирусы.
3. Бактерии.
4. Прионы.
Б. Отметьте отличительные особенности прокариотической клетки:
1. Рибосомы 70S.
2. Наличие пептидогликана в клеточной стенке.
3. Наличие митохондрий.
4. Диплоидный набор генов.
В. Отметьте составные компоненты пептидогликана:
1. Тейхоевые кислоты.
2. N-ацетилглюкозоамин.
3. Липополисарид.
4. Тетрапептид.
Г. Отметьте особенности строения клеточной стенки грамотрицательных бактерий:
1. Мезодиаминопимелиновая кислота.
2. Тейхоевые кислоты.
4. Белки-порины.
Д. Назовите функции спор у бактерий:
1. Сохранение вида.
2. Жароустойчивость.
3. Расселение субстрата.
4. Размножение.
1. Риккетсии.
2. Актиномицеты.
3. Спирохеты.
4. Хламидии.
Ж. Назовите особенности актиномицет:
1. Имеют термолабильные споры.
2. Грамположительные бактерии.
3. Отсутствует клеточная стенка.
4. Имеют извитую форму.
З. Назовите особенности спирохет:
1. Грамотрицательные бактерии.
2. Имеют двигательный фибриллярный аппарат.
3. Имеют извитую форму.
И. Назовите простейшие, обладающие апикальным комплексом, позволяющим проникать внутрь клетки:
1. Малярийный плазмодий.
3. Токсоплазма.
4. Криптоспоридии.
К. Назовите отличительную особенность сложноорганизованных вирусов:
1. Два типа нуклеиновой кислоты.
2. Наличие липидной оболочки.
3. Двойной капсид.
4. Наличие неструктурных белков. Л. Отметьте высшие грибы:
1. Mucor.
2. Candida.
3. Penicillium.
4. Aspergillus.
В бактериальной клетке отсутствует ядро, хромосомы свободно располагаются в цитоплазме. Кроме того, в клетке бактерии отсутствуют мембранные органоиды: митохондрии, ЭПС, аппарат Гольджи и пр. Снаружи клеточная мембрана покрыта клеточной стенкой.



Большинство бактерий передвигаются пассивно, с помощью водных или воздушных течений. Только некоторые из них имеют органеллы движения – жгутики. Жгутики прокариот очень просты по устройству и состоят из белка флагеллина, образующего полый цилиндр диаметром 10–20 нм. Они ввинчиваются в среду, продвигая клетку вперёд. По-видимому, это единственная известная в природе структура, использующая принцип колеса.
По своей форме бактерии делятся на несколько групп:
Кокки (имеют округлую форму);
- бациллы (имеют палочковидную форму);
- спириллы (имеют форму спирали);
- вибрионы (имеют форму запятой).
По способу дыхания бактерии делятся на аэробов (большинство бактерий) и анаэробов (возбудители столбняка, ботулизма, газовой гангрены). Первым для дыхания нужен кислород, для вторых кислород бесполезен или даже ядовит.

Структура прокариотической клетки. Клетка прокариот устроена значительно проще клеток животных и растений. Снаружи она покрыта клеточной стенкой, выполняющей защитные, формирующие и транспортные функции. Жёсткость клеточной стенки обеспечивает муреин. Иногда бактериальная клетка покрыта сверху капсулой или слизистым слоем.
Протоплазма бактерий, как и у эукариот, окружена плазматической мембраной. В мешковидных, трубчатых или пластинчатых впячиваниях мембраны находятся мезосомы, участвующие в процессе дыхания, бактериохлорофилл и другие пигменты.
Генетический материал прокариот не образует ядра, а находится непосредственно в цитоплазме. ДНК бактерий – одиночные кольцевые молекулы, каждая из которых состоит из тысяч и миллионов пар нуклеотидов. Геном бактериальной клетки намного проше, чем у клеток более развитых существ: в среднем ДНК бактерий содержит несколько тысяч генов.
В прокариотических клетках отсутствует эндоплазматическая сеть, а рибосомы свободно плавают в цитоплазме. Нет у прокариот и митохондрий; частично их функции выполняет клеточная мембрана.
Подвижность бактерий обеспечивается жгутиками. Бактерии размножаются путем деления примерно каждые 20 минут (в благоприятных условиях). ДНК реплицируется, каждая дочерняя клетка получает по своей копии родительской ДНК. Возможна также передача ДНК между неделящимися клетками (посредством захвата «голой» ДНК, при помощи бактериофагов или путём конъюгации, когда бактерии соединяются между собой копуляционнымифимбриями), однако увеличения количества особей при этом не происходит. Размножению препятствуют солнечные лучи и продукты их собственной жизнедеятельности.
Поведение бактерий не отличается особой сложностью. Химические рецепторы регистрируют изменения кислотности среды и концентрацию различных веществ: сахаров, аминокислот, кислорода. Многие бактерии реагируют на изменения температуры или освещенности, некоторые бактерии могут чувствовать магнитное поле Земли. При неблагоприятных условиях бактерия покрывается плотной оболочкой, цитоплазма обезвоживается, жизнедеятельность почти прекращается. В таком состоянии споры бактерии могут часами находиться в глубоком вакууме, переносить температуру от –240 °С до +100 °С.