A 19. század első felében. sok kémikus, köztük elsősorban J. von Liebig, fokozatosan arra a következtetésre jutott, hogy a fehérjék a nitrogéntartalmú vegyületek egy speciális osztályát képviselik. A "fehérjék" elnevezés (a görögből.
protos először) G. Mulder holland kémikus javasolta 1840-ben. FIZIKAI TULAJDONSÁGOK A fehérjék szilárd állapotban fehérek, de oldatban színtelenek, hacsak nem hordoznak valamilyen kromofor (színes) csoportot, például hemoglobint. A vízben való oldhatóság nagyon eltérő a különböző fehérjék között. Változik a pH-tól és az oldatban lévő sók koncentrációjától is, így ki lehet választani azokat a körülményeket, amelyek mellett az egyik fehérje szelektíven kicsapódik más fehérjék jelenlétében. Ezt a "kisózási" módszert széles körben használják fehérjék izolálására és tisztítására. A tisztított fehérje gyakran kristályok formájában válik ki az oldatból.Más vegyületekhez képest a fehérjék molekulatömege nagyon nagy, több ezertől sok millió daltonig terjed. Ezért az ultracentrifugálás során a fehérjék ülepednek, és különböző sebességgel. A fehérjemolekulákban található pozitív és negatív töltésű csoportok miatt eltérő sebességgel és elektromos térben mozognak. Ez az elektroforézis alapja, amely módszer az egyes fehérjék komplex keverékekből történő izolálására szolgál. A fehérjéket kromatográfiával is tisztítják.
KÉMIAI TULAJDONSÁGOK Szerkezet. A fehérjék polimerek, pl. ismétlődő monomer egységekből vagy alegységekből láncokként felépülő molekulák, amelyek szerepét töltik be a -aminosavak. Az aminosavak általános képlete ahol R hidrogénatom vagy valamilyen szerves csoport.Egy fehérje molekula (polipeptid lánc) csak viszonylag kis számú aminosavból vagy több ezer monomer egységből állhat. Az aminosavak láncban való kombinációja azért lehetséges, mert mindegyiknek két különböző kémiai csoportja van: egy bázikus tulajdonságokkal rendelkező aminocsoport,
NH 2 és egy savas karboxilcsoport, COOH. Mindkét csoport kapcsolatban áll a -szénatom. Egy aminosav karboxilcsoportja amid (peptid) kötést képezhet egy másik aminosav aminocsoportjával: Két aminosav ily módon történő összekapcsolása után a lánc meghosszabbítható, ha a második aminosavhoz egy harmadikat adunk, és így tovább. Amint az a fenti egyenletből látható, amikor egy peptidkötés képződik, egy vízmolekula szabadul fel. Savak, lúgok vagy proteolitikus enzimek jelenlétében a reakció az ellenkező irányba megy végbe: a polipeptidlánc víz hozzáadásával aminosavakra hasad. Ezt a reakciót hidrolízisnek nevezik. A hidrolízis spontán megy végbe, és energiára van szükség ahhoz, hogy az aminosavakat polipeptidláncba kapcsolják.
Két aminosav ily módon történő összekapcsolása után a lánc meghosszabbítható, ha a második aminosavhoz egy harmadikat adunk, és így tovább. Amint az a fenti egyenletből látható, amikor egy peptidkötés képződik, egy vízmolekula szabadul fel. Savak, lúgok vagy proteolitikus enzimek jelenlétében a reakció az ellenkező irányba megy végbe: a polipeptidlánc víz hozzáadásával aminosavakra hasad. Ezt a reakciót hidrolízisnek nevezik. A hidrolízis spontán megy végbe, és energiára van szükség ahhoz, hogy az aminosavakat polipeptidláncba kapcsolják. Egy karboxilcsoport és egy amidcsoport (vagy a prolin aminosavnál hasonló imidcsoport) minden aminosavban megtalálható, de az aminosavak közötti különbséget a csoport, vagyis az „oldallánc” jellege határozza meg. amelyet fent a levél jelez
R . Az oldallánc szerepét betöltheti egy hidrogénatom, mint például a glicin aminosavban, vagy valamilyen terjedelmes csoport, mint a hisztidinben és a triptofánban. Egyes oldalláncok kémiailag inertek, míg mások kifejezetten reaktívak.Sok ezer különböző aminosav szintetizálható, és sokféle aminosav fordul elő a természetben, de a fehérjeszintézishez csak 20 féle aminosavat használnak: alanin, arginin, aszparagin, aszparaginsav, valin, hisztidin, glicin, glutamin, glutaminsav sav, izoleucin, leucin, lizin, metionin, prolin, szerin, tirozin, treonin, triptofán, fenilalanin és cisztein (a fehérjékben a cisztein dimerként is jelen lehet
cisztin). Igaz, egyes fehérjék a rendszeresen előforduló húsz aminosav mellett más aminosavakat is tartalmaznak, de ezek a felsorolt húsz közül valamelyiknek a fehérjébe kerülése utáni módosítása következtében jönnek létre.Optikai tevékenység. A glicin kivételével minden aminosav rendelkezik a A -szénatomhoz négy különböző csoport kapcsolódik. Geometriai szempontból négy különböző csoport kétféleképpen kapcsolható össze, ennek megfelelően két konfiguráció, vagy két izomer lehetséges, amelyek úgy viszonyulnak egymáshoz, mint egy tárgy a tükörképéhez, azaz. mint a bal kéz jobbra. Az egyik konfigurációt balosnak vagy balkezesnek ( L ), a másik jobb vagy jobbra forgató ( D ), mivel két ilyen izomer különbözik a polarizált fény síkjának forgási irányában. Csak a fehérjékben található L -aminosavak (kivétel a glicin; csak egy formában ábrázolható, mivel négy csoportjából kettő azonos), és mindegyik optikailag aktív (mivel csak egy izomer van). D -az aminosavak ritkák a természetben; egyes antibiotikumokban és a baktériumok sejtfalában találhatók.Aminosav szekvencia. A polipeptidláncban az aminosavak nem véletlenszerűen, hanem meghatározott sorrendben helyezkednek el, és ez a sorrend határozza meg a fehérje funkcióit és tulajdonságait. A 20 fajta aminosav sorrendjének változtatásával rengeteg különböző fehérjét hozhatunk létre, ahogyan az ábécé betűiből is sokféle szöveget készíthetünk.A múltban egy fehérje aminosavszekvenciájának meghatározása gyakran több évig tartott. A közvetlen meghatározás továbbra is meglehetősen munkaigényes feladat, bár készültek olyan eszközök, amelyek lehetővé teszik ennek automatikus végrehajtását. Általában könnyebb meghatározni a megfelelő gén nukleotidszekvenciáját, és abból következtetni a fehérje aminosavsorrendjére. A mai napig sok száz fehérje aminosavszekvenciáját határozták meg már. A megfejtett fehérjék funkciói általában ismertek, és ez segít elképzelni a hasonló, például rosszindulatú daganatokban képződött fehérjék lehetséges funkcióit.
Komplex fehérjék. A csak aminosavakból álló fehérjéket egyszerűnek nevezzük. Gyakran azonban a polipeptidlánchoz fématom vagy valamilyen kémiai vegyület kapcsolódik, amely nem aminosav. Az ilyen fehérjéket komplexnek nevezzük. Példa erre a hemoglobin: vas-porfirint tartalmaz, amely meghatározza vörös színét, és lehetővé teszi, hogy oxigénhordozóként működjön.A legtöbb összetett fehérje neve a kapcsolódó csoportok jellegét jelzi: a glikoproteinek cukrokat, a lipoproteinek zsírokat tartalmaznak. Ha egy enzim katalitikus aktivitása a kapcsolódó csoporttól függ, akkor azt protetikus csoportnak nevezzük. Gyakran egy vitamin egy protéziscsoport szerepét tölti be, vagy annak része. Az A-vitamin például, amely a retina egyik fehérjéhez kapcsolódik, meghatározza annak fényérzékenységét.
Harmadlagos szerkezet. Nem annyira magának a fehérjének (az elsődleges szerkezetnek) az aminosavsorrendje a fontos, hanem a térben való elhelyezkedése. A hidrogénionok a polipeptidlánc teljes hosszában szabályos hidrogénkötéseket alkotnak, amelyek hélix vagy réteg alakját adják (másodlagos szerkezet). Az ilyen hélixek és rétegek kombinációjából a következő rend kompakt formája alakul ki: a fehérje harmadlagos szerkezete. A lánc monomer egységeit tartó kötések körül kis szögű elfordulások lehetségesek. Ezért tisztán geometriai szempontból bármely polipeptidlánc lehetséges konfigurációinak száma végtelenül nagy. A valóságban minden fehérje általában csak egy konfigurációban létezik, amelyet az aminosavszekvenciája határoz meg. Ez a szerkezet nem merev, olyan, mintha « lélegzik” egy bizonyos átlagos konfiguráció körül ingadozik. Az áramkör olyan konfigurációba van összehajtva, amelyben a szabad energia (munkatermelő képesség) minimális, mint ahogy a felszabaduló rugó is csak a minimális szabad energiának megfelelő állapotba szorítja össze. Gyakran a lánc egyik része mereven kapcsolódik a másikhoz diszulfiddal ( SS) kötések két ciszteinmaradék között. Részben ezért játszik különösen fontos szerepet a cisztein az aminosavak között.A fehérjék szerkezetének összetettsége olyan nagy, hogy még nem lehet kiszámítani egy fehérje harmadlagos szerkezetét, még akkor sem, ha ismert az aminosavsorrendje. De ha lehetséges fehérjekristályokat előállítani, akkor annak harmadlagos szerkezete röntgendiffrakcióval meghatározható.
Strukturális, kontraktilis és néhány más fehérjében a láncok megnyúltak, és a közelben elhelyezkedő enyhén hajtogatott láncok rostokat képeznek; a rostok pedig nagyobb rostképződményekké gyűrődnek. Az oldatban lévő fehérjék többsége azonban gömb alakú: a láncok gömbölyűbe vannak tekercselve, mint a fonal egy golyóban. A szabad energia ezzel a konfigurációval minimális, mivel a hidrofób („víztaszító”) aminosavak a gömböcskében vannak elrejtve, és hidrofil („vízvonzó”) aminosavak találhatók a felszínén.
Sok fehérje több polipeptidlánc komplexe. Ezt a szerkezetet a fehérje kvaterner szerkezetének nevezik. A hemoglobin molekula például négy alegységből áll, amelyek mindegyike globuláris fehérje.
A strukturális fehérjék lineáris konfigurációjukból adódóan nagyon nagy szakítószilárdságú rostokat képeznek, míg a globuláris konfiguráció lehetővé teszi, hogy a fehérjék specifikus kölcsönhatásba lépjenek más vegyületekkel. A golyók felületén, ha a láncok megfelelően vannak elhelyezve, bizonyos alakú üregek jelennek meg, amelyekben reaktív kémiai csoportok találhatók. Ha egy adott fehérje egy enzim, akkor valamilyen anyag egy másik, általában kisebb molekulája kerül be egy ilyen üregbe, ahogy a kulcs a zárba; ilyenkor az üregben elhelyezkedő kémiai csoportok hatására megváltozik a molekula elektronfelhőjének konfigurációja, és ez bizonyos reakcióra kényszeríti. Ily módon az enzim katalizálja a reakciót. Az antitest molekulákban is vannak üregek, amelyekben különféle idegen anyagok kötődnek, és ezáltal ártalmatlanná válnak. A „zár és kulcs” modell, amely a fehérjék más vegyületekkel való kölcsönhatását magyarázza, lehetővé teszi, hogy megértsük az enzimek és antitestek specifitását, i. csak bizonyos vegyületekkel reagálnak.
Fehérjék különböző típusú szervezetekben. A különböző növény- és állatfajokban ugyanazt a funkciót betöltő, ezért ugyanazt a nevet viselő fehérjék szintén hasonló konfigurációval rendelkeznek. Aminosavszekvenciájukban azonban némileg eltérnek. Mivel a fajok eltérnek egy közös őstől, bizonyos pozíciókban egyes aminosavakat mutációk helyettesítenek másokkal. Az örökletes betegségeket okozó káros mutációkat a természetes szelekció eliminálja, de a jótékony vagy legalább semleges mutációk fennmaradhatnak. Minél közelebb van egymáshoz a két faj, annál kisebb különbségek találhatók a fehérjékben.Egyes fehérjék viszonylag gyorsan változnak, mások nagyon konzerváltak. Ez utóbbihoz tartozik például a citokróm Val vel légzőszervi enzim, amely a legtöbb élő szervezetben megtalálható. Emberben és csimpánzban aminosav-szekvenciája azonos, a citokrómban Val vel A búzában az aminosavak mindössze 38%-a különbözött egymástól. Még az embert és a baktériumokat is összehasonlítva a citokrómok hasonlósága Val vel(a különbségek itt az aminosavak 65%-át érintik) még mindig látható, bár a baktériumok és az emberek közös őse körülbelül kétmilliárd évvel ezelőtt élt a Földön. Manapság az aminosav-szekvenciák összehasonlítását gyakran használják filogenetikai (családi) fa megalkotására, amely tükrözi a különböző szervezetek közötti evolúciós kapcsolatokat.
Denaturáció. A szintetizált fehérjemolekula, a folding, elnyeri jellegzetes konfigurációját. Ez a konfiguráció azonban tönkretehető hevítéssel, pH-érték megváltoztatásával, szerves oldószerekkel való érintkezéssel, sőt, az oldat egyszerű rázásával, amíg buborékok nem jelennek meg a felületén. Az így módosított fehérjét denaturáltnak nevezzük; elveszti biológiai aktivitását és általában oldhatatlanná válik. A denaturált fehérjék jól ismert példái a főtt tojás vagy a tejszínhab. A mindössze mintegy száz aminosavat tartalmazó kisméretű fehérjék képesek renaturálódni, azaz. visszaszerzi az eredeti konfigurációt. De a legtöbb fehérje egyszerűen kusza polipeptidláncok tömegévé változik, és nem állítja vissza korábbi konfigurációját.Az aktív fehérjék izolálásának egyik fő nehézsége a denaturációval szembeni rendkívüli érzékenységük. A fehérjéknek ez a tulajdonsága hasznos az élelmiszerek tartósításában: a magas hőmérséklet visszafordíthatatlanul denaturálja a mikroorganizmusok enzimjeit, és a mikroorganizmusok elpusztulnak.
PROTEIN SZINTÉZIS A fehérje szintéziséhez az élő szervezetnek rendelkeznie kell egy olyan enzimrendszerrel, amely képes az egyik aminosavat a másikhoz kapcsolni. Információforrásra is szükség van annak meghatározásához, hogy mely aminosavakat érdemes kombinálni. Mivel a szervezetben több ezerféle fehérje található, és mindegyik átlagosan több száz aminosavból áll, a szükséges információnak valóban óriásinak kell lennie. A géneket alkotó nukleinsavmolekulákban tárolódik (hasonlóan ahhoz, ahogy egy felvételt mágnesszalagon tárolnak). Cm . ÖRÖKLETES is; NUKLEINSAVAK.Enzimaktiválás. Az aminosavakból szintetizált polipeptidlánc nem mindig fehérje a végső formájában. Sok enzimet először inaktív prekurzorként szintetizálnak, és csak azután válnak aktívvá, hogy egy másik enzim eltávolít több aminosavat a lánc egyik végén. Az emésztőenzimek egy része, mint például a tripszin, ebben az inaktív formában szintetizálódik; ezek az enzimek aktiválódnak az emésztőrendszerben a lánc terminális fragmentumának eltávolítása következtében. Az inzulin hormon, melynek molekulája aktív formájában két rövid láncból áll, egy lánc formájában szintetizálódik, az ún. proinzulin. Ennek a láncnak a középső részét ezután eltávolítják, és a fennmaradó fragmensek összekapcsolódnak, és létrehozzák az aktív hormonmolekulát. Komplex fehérjék csak azután jönnek létre, hogy a fehérjéhez egy meghatározott kémiai csoport kapcsolódik, és ehhez a kötődéshez gyakran enzim is szükséges.Metabolikus keringés. A szén, nitrogén vagy hidrogén radioaktív izotópjaival jelölt aminosavak etetése után a címke gyorsan beépül a fehérjékbe. Ha a jelölt aminosavak már nem jutnak be a szervezetbe, a fehérjékben lévő címkék mennyisége csökkenni kezd. Ezek a kísérletek azt mutatják, hogy a keletkező fehérjék életük végéig nem maradnak meg a szervezetben. Néhány kivételtől eltekintve mindegyik dinamikus állapotban van, folyamatosan aminosavakra bomlik, majd újra szintetizálódnak.Néhány fehérje lebomlik, amikor a sejtek elpusztulnak, és elpusztulnak. Ez mindig megtörténik például a bél belső felületét borító vörösvértestekkel és hámsejtekkel. Ezenkívül a fehérjék lebontása és újraszintézise az élő sejtekben is megtörténik. Furcsa módon kevesebbet tudunk a fehérjék lebontásáról, mint a szintézisükről. Nyilvánvaló azonban, hogy a lebontásban olyan proteolitikus enzimek vesznek részt, amelyek hasonlóak azokhoz, amelyek a fehérjéket aminosavakra bontják az emésztőrendszerben.
A különböző fehérjék felezési ideje több órától több hónapig terjed. Az egyetlen kivétel a kollagén molekula. Kialakulása után stabilak maradnak, és nem újulnak meg vagy cserélnek ki. Idővel azonban bizonyos tulajdonságaik megváltoznak, különösen a rugalmasságuk, és mivel nem újulnak meg, ez bizonyos életkorral összefüggő változásokat eredményez, például ráncok megjelenését a bőrön.
Szintetikus fehérjék. A kémikusok már régóta megtanulták az aminosavak polimerizálását, de az aminosavakat szabálytalanul kombinálják, így az ilyen polimerizáció termékei alig hasonlítanak a természetesekhez. Igaz, lehetséges az aminosavak adott sorrendben történő kombinálása, ami lehetővé teszi néhány biológiailag aktív fehérje, különösen az inzulin előállítását. A folyamat meglehetősen bonyolult, és így csak azokat a fehérjéket lehet előállítani, amelyek molekulái körülbelül száz aminosavat tartalmaznak. Ehelyett előnyösebb a kívánt aminosavszekvenciának megfelelő gén nukleotidszekvenciáját szintetizálni vagy izolálni, majd ezt a gént bevinni egy baktériumba, amely replikáció útján nagy mennyiségben termeli a kívánt terméket. Ennek a módszernek azonban vannak hátrányai is. Cm . GÉNÉRZET is. FEHÉRJE ÉS TÁPLÁLKOZÁS Amikor a fehérjéket a szervezetben aminosavakra bontják, ezek az aminosavak újra felhasználhatók fehérjék szintézisére. Ugyanakkor maguk az aminosavak is lebomlanak, így nem hasznosulnak teljesen újra. Az is világos, hogy a növekedés, a terhesség és a sebgyógyulás során a fehérjeszintézisnek meg kell haladnia a lebontást. A szervezet folyamatosan veszít bizonyos fehérjékből; Ezek a haj, a köröm és a bőr felszíni rétegének fehérjéi. Ezért a fehérjék szintéziséhez minden szervezetnek aminosavakat kell kapnia az élelmiszerből. A zöld növények CO-ból szintetizálódnak 2 , a víz és az ammónia vagy a nitrátok mind a 20 aminosav, amelyek a fehérjékben találhatók. Sok baktérium képes aminosavak szintetizálására is cukor (vagy ennek megfelelője) és rögzített nitrogén jelenlétében, de a cukrot végső soron zöld növények szállítják. Az állatok korlátozott mértékben képesek aminosavakat szintetizálni; zöld növények vagy más állatok fogyasztásával nyernek aminosavakat. Az emésztőrendszerben a felszívódott fehérjék aminosavakra bomlanak le, ez utóbbiak felszívódnak, és belőlük épülnek fel az adott szervezetre jellemző fehérjék. A felszívódott fehérjék egyike sem épül be a testszerkezetekbe. Az egyetlen kivétel az, hogy sok emlősben egyes anyai antitestek épségben a méhlepényen keresztül a magzati vérkeringésbe juthatnak, és az anyatejen (különösen kérődzőknél) közvetlenül a születés után átjuthatnak az újszülöttbe.Fehérje szükséglet. Nyilvánvaló, hogy az élet fenntartásához a szervezetnek bizonyos mennyiségű fehérjét kell kapnia az élelmiszerből. Ennek az igénynek a mértéke azonban számos tényezőtől függ. A szervezetnek szüksége van táplálékra energiaforrásként (kalória) és szerkezeti felépítéséhez. Az energiaigény az első. Ez azt jelenti, hogy amikor az étrendben kevés a szénhidrát és a zsír, az étkezési fehérjéket nem saját fehérjék szintézisére, hanem kalóriaforrásként használják fel. A hosszan tartó koplalás során még a saját fehérjéit is felhasználják az energiaszükségletek kielégítésére. Ha elegendő szénhidrát van az étrendben, akkor a fehérjefogyasztás csökkenthető.Nitrogén egyensúly. Átlagosan kb. A fehérje teljes tömegének 16%-a nitrogén. A fehérjékben található aminosavak lebontásakor a bennük lévő nitrogén a vizelettel és (kisebb mértékben) a széklettel ürül ki a szervezetből különböző nitrogéntartalmú vegyületek formájában. Ezért célszerű olyan mutatót használni, mint a nitrogénegyensúly a fehérjetáplálkozás minőségének értékelésére, pl. a szervezetbe jutó nitrogén mennyisége és a naponta kiválasztott nitrogén mennyisége közötti különbség (grammban). Normál táplálkozás mellett felnőtteknél ezek a mennyiségek egyenlőek. Egy növekvő szervezetben a kiválasztott nitrogén mennyisége kisebb, mint a bevitt mennyiség, azaz. az egyenleg pozitív. Ha fehérjehiány van az étrendben, az egyenleg negatív. Ha az étrendben van elegendő kalória, de nincs benne fehérje, a szervezet megtakarítja a fehérjéket. Ezzel párhuzamosan a fehérjeanyagcsere lelassul, az aminosavak ismételt hasznosítása a fehérjeszintézisben a lehető legnagyobb hatékonysággal történik. A veszteségek azonban elkerülhetetlenek, és a nitrogéntartalmú vegyületek továbbra is a vizelettel és részben a széklettel választódnak ki. A fehérjeböjt során a szervezetből naponta kiürülő nitrogén mennyisége a napi fehérjehiány mértékeként szolgálhat. Természetes az a feltételezés, hogy az étrendbe e hiánynak megfelelő mennyiségű fehérje bevezetésével a nitrogén-egyensúly helyreállítható. Azonban nem. Miután megkapta ezt a fehérjemennyiséget, a szervezet kevésbé hatékonyan kezdi el felhasználni az aminosavakat, ezért további fehérjére van szükség a nitrogén egyensúly helyreállításához.Ha az étrendben lévő fehérje mennyisége meghaladja a nitrogén egyensúly fenntartásához szükséges mennyiséget, úgy tűnik, nincs baj. A felesleges aminosavakat egyszerűen energiaforrásként használják fel. Különösen szembetűnő példa, hogy az eszkimók kevés szénhidrátot és körülbelül tízszer annyi fehérjét fogyasztanak, mint ami a nitrogén egyensúly fenntartásához szükséges. A legtöbb esetben azonban a fehérje energiaforrásként való felhasználása nem előnyös, mert adott mennyiségű szénhidrát sokkal több kalóriát tud termelni, mint ugyanannyi fehérje. A szegény országokban az emberek szénhidrátokból szerzik be kalóriájukat, és minimális mennyiségű fehérjét fogyasztanak.
Ha a szervezet a szükséges mennyiségű kalóriát nem fehérje termékek formájában kapja meg, akkor a nitrogén egyensúly fenntartását biztosító minimális fehérje mennyiség kb. 30 g naponta. Körülbelül ennyi fehérjét tartalmaz négy szelet kenyér vagy 0,5 liter tej. Általában valamivel nagyobb számot tartanak optimálisnak; 50-70 g ajánlott.
Esszenciális aminosavak. Eddig a fehérjét egésznek tekintették. Eközben ahhoz, hogy a fehérjeszintézis megtörténjen, minden szükséges aminosavnak jelen kell lennie a szervezetben. Az állat szervezete maga is képes szintetizálni az aminosavak egy részét. Cserélhetőnek nevezik őket, mert nem feltétlenül kell jelen lenniük az étrendben, csak az a fontos, hogy a fehérje, mint nitrogénforrás összességében elegendő legyen; majd ha hiány van a nem esszenciális aminosavakban, akkor a szervezet képes ezeket szintetizálni a feleslegben lévők rovására. A fennmaradó, „esszenciális” aminosavakat nem lehet szintetizálni, táplálékkal kell bejuttatni a szervezetbe. Az emberek számára nélkülözhetetlenek a valin, leucin, izoleucin, treonin, metionin, fenilalanin, triptofán, hisztidin, lizin és arginin. (Bár az arginin szintetizálható a szervezetben, az esszenciális aminosavak közé sorolják, mert újszülöttekben és felnövő gyermekekben nem termelődik kellő mennyiségben. Másrészt ezeknek az aminosavak egy része az élelmiszerből feleslegessé válhat egy felnőtt számára személy.)Az esszenciális aminosavak listája megközelítőleg megegyezik más gerinceseknél, sőt rovaroknál is. A fehérjék tápértékét általában úgy határozzák meg, hogy növekvő patkányokkal etetik, és figyelemmel kísérik az állatok súlygyarapodását.
A fehérjék tápértéke. A fehérje tápértékét a leginkább hiányzó esszenciális aminosav határozza meg. Illusztráljuk ezt egy példával. A szervezetünkben lévő fehérjék átlagosan kb. 2 tömeg% triptofán. Tegyük fel, hogy az étrendben 10 g 1% triptofánt tartalmazó fehérje van, és van benne elegendő egyéb esszenciális aminosav. Esetünkben ebből a nem teljes fehérjéből 10 g lényegében 5 g teljes fehérjével egyenértékű; a maradék 5 g csak energiaforrásul szolgálhat. Megjegyzendő, hogy mivel az aminosavak gyakorlatilag nem raktározódnak a szervezetben, és a fehérjeszintézis létrejöttéhez az összes aminosavnak egyszerre kell jelen lennie, az esszenciális aminosavak bevitelének hatása csak akkor mutatható ki, ha az összes egyszerre jutnak be a szervezetbe. A legtöbb állati fehérje átlagos összetétele megközelíti az emberi szervezet fehérje átlagos összetételét, így nem valószínű, hogy aminosavhiánnyal szembesülünk, ha étrendünk gazdag élelmiszerekben, például húsban, tojásban, tejben és sajtban. Vannak azonban olyan fehérjék, mint például a zselatin (a kollagén denaturációjának terméke), amelyek nagyon kevés esszenciális aminosavat tartalmaznak. A növényi fehérjék, bár ebben az értelemben jobbak, mint a zselatin, esszenciális aminosavakban is szegények; Különösen alacsony a lizin- és triptofántartalmuk. Ennek ellenére a tisztán vegetáriánus étrend egyáltalán nem tekinthető károsnak, hacsak nem fogyaszt valamivel nagyobb mennyiségű növényi fehérjét, ami elegendő ahhoz, hogy a szervezetet esszenciális aminosavakkal láthassa el. A növények magjaiban tartalmazzák a legtöbb fehérjét, különösen a búza és a különféle hüvelyesek magjaiban. A fiatal hajtások, például a spárga, szintén gazdagok fehérjében.Szintetikus fehérjék az étrendben. Kis mennyiségben szintetikus esszenciális aminosavak vagy aminosavban gazdag fehérjék hozzáadásával a nem teljes fehérjékhez, például a kukoricafehérjékhez, ez utóbbiak tápértéke jelentősen növelhető, i.e. ezzel növelve az elfogyasztott fehérje mennyiségét. Egy másik lehetőség a baktériumok vagy élesztők tenyésztése kőolaj-szénhidrogéneken nitrogénforrásként nitrátok vagy ammónia hozzáadásával. Az így nyert mikrobiális fehérje szolgálhat baromfi vagy haszonállat takarmányaként, vagy közvetlenül fogyasztható az ember számára. A harmadik, széles körben alkalmazott módszer a kérődzők fiziológiáját használja fel. Kérődzőknél a gyomor kezdeti részében az ún. A bendőben a baktériumok és a protozoonok speciális formái élnek, amelyek a nem teljes növényi fehérjéket teljesebb mikrobiális fehérjékké alakítják, ezek pedig az emésztés és felszívódás után állati fehérjékké alakulnak. A karbamid, egy olcsó szintetikus nitrogéntartalmú vegyület, adható az állati takarmányokhoz. A bendőben élő mikroorganizmusok karbamid-nitrogént használnak a szénhidrátok (amiből sokkal több van a takarmányban) fehérjévé alakítására. Az állati takarmányban lévő összes nitrogén körülbelül egyharmada származhat karbamid formájában, ami lényegében bizonyos mértékig a fehérje kémiai szintézisét jelenti. Az USA-ban ez a módszer fontos szerepet játszik a fehérje beszerzésének egyik módjaként.IRODALOM Murray R., Grenner D., Mayes P., Rodwell W. Az emberi biokémia, vol. 12. M., 1993Alberts B, Bray D, Lewis J és mtsai. Molekuláris sejtbiológia, vol. 13. M., 1994
2. jegy. 1. A lipid jellegű alapvető táplálkozási tényezők. Egyes lipidek nem szintetizálódnak az emberi szervezetben, ezért alapvető táplálkozási tényezők. Ide tartoznak a két vagy több kettős kötést tartalmazó zsírsavak (polién) - esszenciális zsírsavak. Ezen savak egy része a helyi hormonok – eikozanoidok – szintézisének szubsztrátja (8.10. téma).
Zsírban oldódó vitaminok különböző funkciókat lát el: A vitamin részt vesz a látás, valamint a sejtnövekedés és differenciálódás folyamatában; bizonyítottan képes bizonyos típusú daganatok növekedését gátolni; K vitamin részt vesz a véralvadásban; D-vitamin részt vesz a kalcium-anyagcsere szabályozásában; E vitamin- antioxidáns, gátolja a szabad gyökök képződését és így ellensúlyozza a lipidperoxidáció következtében fellépő sejtkárosodást.
Dokumentum
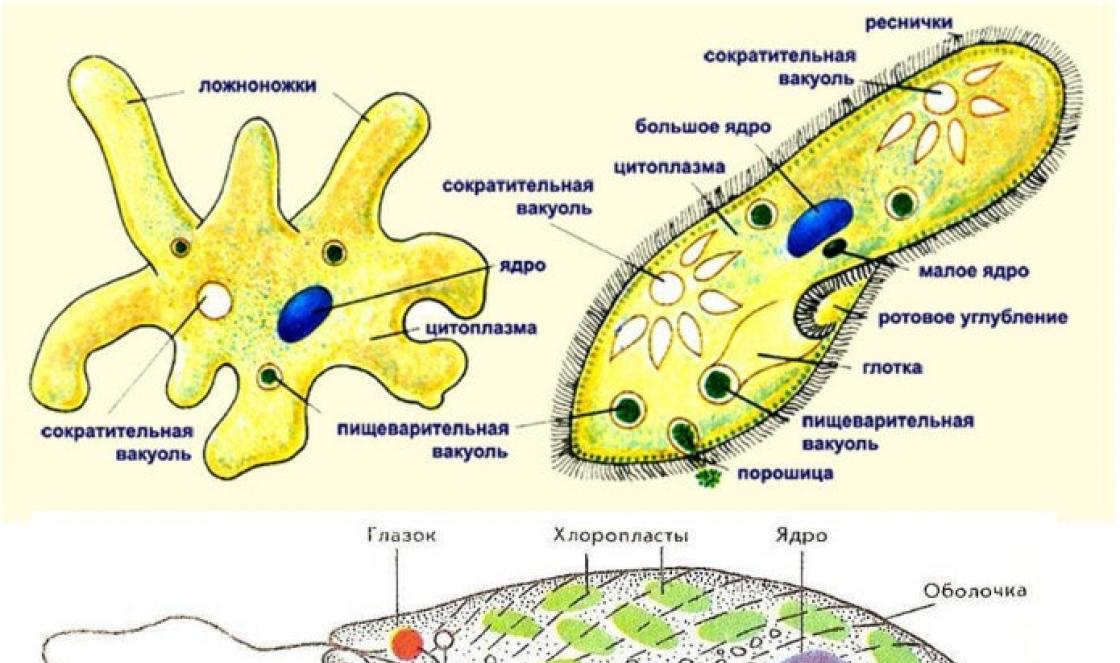
2. A fehérjék szerkezete és szerkezeti szerveződési szintjei
A fehérjék szerkezeti szerveződésének négy szintje van: primer, szekunder, tercier és kvaterner. Minden szintnek megvannak a maga sajátosságai.
Elsődleges fehérje szerkezet
A fehérjék elsődleges szerkezete aminosavakból álló lineáris polipeptidlánc, amelyet peptidkötések kapcsolnak össze. Az elsődleges szerkezet a fehérjemolekula szerkezeti szerveződésének legegyszerűbb szintje. Nagy stabilitást az egyik aminosav α-aminocsoportja és egy másik aminosav α-karboxilcsoportja közötti kovalens peptidkötések biztosítanak. [előadás].
Ha a prolin vagy a hidroxiprolin iminocsoportja részt vesz a peptidkötés kialakításában, akkor annak más formája van [előadás].
Amikor peptidkötések képződnek a sejtekben, először az egyik aminosav karboxilcsoportja aktiválódik, majd egyesül egy másik aminocsoportjával. A polipeptidek laboratóriumi szintézisét megközelítőleg hasonló módon végezzük.
A peptidkötés egy polipeptidlánc ismétlődő fragmentuma. Számos olyan tulajdonsággal rendelkezik, amelyek nemcsak az elsődleges szerkezet alakját, hanem a polipeptidlánc magasabb szintű szerveződését is befolyásolják:

koplanaritás - a peptidcsoportba tartozó összes atom ugyanabban a síkban van;
két rezonanciaformában való létezés képessége (keto vagy enol forma);
a szubsztituensek transz-helyzete a C-N kötéshez képest;
hidrogénkötések kialakításának képessége, és mindegyik peptidcsoport két hidrogénkötést tud kialakítani más csoportokkal, beleértve a peptidcsoportokat is.
Kivételt képeznek a prolin vagy hidroxiprolin aminocsoportját tartalmazó peptidcsoportok. Csak egy hidrogénkötést képesek létrehozni (lásd fent). Ez befolyásolja a fehérje másodlagos szerkezetének kialakulását. A polipeptidlánc azon a területen, ahol a prolin vagy a hidroxiprolin található, könnyen meghajlik, mivel a szokásos módon nem tartja egy második hidrogénkötésben.
A peptidek és polipeptidek nómenklatúrája. A peptidek neve az őket alkotó aminosavak nevéből áll. Két aminosav alkot egy dipeptidet, három egy tripeptidet, négy egy tetrapeptidet stb. Minden peptid vagy polipeptid lánc tetszőleges hosszúságú tartalmaz egy N-terminális aminosavat, amely szabad aminocsoportot és egy C-terminális aminosavat tartalmaz szabad karboxilcsoportot csoport. A polipeptidek elnevezésekor az összes aminosavat egymás után felsoroljuk, az N-terminálistól kezdve, a nevükben a C-terminális kivételével az -in utótagot -yl-lel helyettesítve (mivel a peptidekben lévő aminosavak már nem rendelkeznek karboxilcsoport, hanem karbonilcsoport). Például az ábrán látható név. 1 tripeptid - leuc iszap fenilalan iszap treon ban ben.
A fehérje elsődleges szerkezetének jellemzői. A polipeptid lánc gerincében merev struktúrák (lapos peptidcsoportok) váltakoznak a viszonylag mobil régiókkal (-CHR), amelyek képesek a kötések körül forogni. A polipeptidlánc ilyen szerkezeti jellemzői befolyásolják annak térbeli elrendezését.
A fehérje másodlagos szerkezete
A másodlagos szerkezet a polipeptidlánc rendezett szerkezetté való hajtogatásának módja az azonos lánc peptidcsoportjai vagy a szomszédos polipeptidláncok közötti hidrogénkötések kialakulása miatt. Konfigurációjuk szerint a másodlagos struktúrákat helikális (α-hélix) és rétegesen hajtogatott (β-szerkezet és kereszt-β-forma) részekre osztják.


α-Hélix. Ez egy olyan típusú másodlagos fehérjeszerkezet, amely szabályos hélixnek tűnik, és az egyik polipeptidláncon belüli interpeptid hidrogénkötések következtében jön létre. Az α-hélix szerkezetének modelljét (2. ábra), amely figyelembe veszi a peptidkötés összes tulajdonságát, Pauling és Corey javasolta. Az α-hélix főbb jellemzői:
a polipeptid lánc helikális konfigurációja helikális szimmetriával;
hidrogénkötések kialakítása az egyes első és negyedik aminosavmaradékok peptidcsoportjai között;
a spirális fordulatok szabályossága;
az α-hélixben lévő összes aminosav ekvivalenciája, függetlenül az oldalgyökök szerkezetétől;
aminosavak oldalgyökei nem vesznek részt az α-hélix kialakulásában.
Külsőleg az α-hélix úgy néz ki, mint egy elektromos tűzhely enyhén megfeszített spirálja. Az első és negyedik peptidcsoport közötti hidrogénkötések szabályossága meghatározza a polipeptid lánc fordulatainak szabályosságát. Egy fordulat magassága vagy az α-hélix osztásköze 0,54 nm; 3,6 aminosavból áll, azaz minden aminosav a tengely mentén (egy aminosav magassága) 0,15 nm-rel mozog (0,54:3,6 = 0,15 nm), ami lehetővé teszi, hogy az összes aminosav ekvivalenciájáról beszéljünk az α-hélixben. Az α-hélix szabályossági periódusa 5 fordulat vagy 18 aminosav; egy periódus hossza 2,7 nm. Rizs. 3. Pauling-Corey a-helix modell
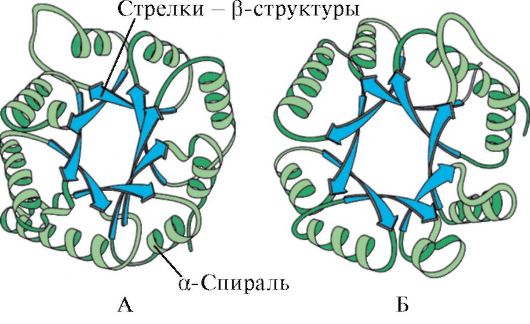
β-Struktúra. Ez a másodlagos szerkezet olyan típusa, amelynek a polipeptidlánc enyhén ívelt konfigurációja van, és az egyik polipeptidlánc vagy a szomszédos polipeptidláncok egyes szakaszain belüli interpeptid hidrogénkötések alkotják. Réteges redős szerkezetnek is nevezik. A β-struktúráknak különféle fajtái vannak. A fehérje egy polipeptidlánca által alkotott korlátozott rétegű régiókat kereszt-β formának (rövid β szerkezet) nevezzük. A hidrogénkötések kereszt-β formában jönnek létre a polipeptidlánc hurkainak peptidcsoportjai között. Egy másik típus - a teljes β-struktúra - a teljes polipeptidláncra jellemző, amely hosszúkás alakú, és a szomszédos párhuzamos polipeptidláncok között interpeptid hidrogénkötések tartják fenn (3. ábra). Ez a szerkezet egy harmonika fújtatójára hasonlít. Sőt, a β-struktúrák variánsai is lehetségesek: létrejöhetnek párhuzamos láncokkal (a polipeptidláncok N-terminális végei ugyanabba az irányba vannak irányítva) és antiparallel módon (az N-terminális végek különböző irányokba vannak irányítva). Az egyik réteg oldalgyökei egy másik réteg oldalgyökei közé kerülnek.
A fehérjékben a hidrogénkötések átrendeződése miatt lehetséges az α-struktúrákból a β-struktúrákba és vissza. A lánc mentén kialakuló szabályos interpeptid hidrogénkötések helyett (aminek köszönhetően a polipeptidlánc spirálba csavarodik) a spirális szakaszok feltekernek, és a hidrogénkötések bezáródnak a polipeptidláncok megnyúlt fragmentumai közé. Ez az átmenet a keratinban, a haj fehérjéjében található. Lúgos mosószerrel történő hajmosáskor a β-keratin spirális szerkezete könnyen tönkremegy, és α-keratinná alakul (a göndör haj kiegyenesedik).
A fehérjék szabályos másodlagos struktúráinak (α-hélixek és β-struktúrák) elpusztítását a kristályok megolvadásával analóg módon polipeptidek „olvadásának” nevezik. Ebben az esetben a hidrogénkötések megszakadnak, és a polipeptid láncok véletlenszerű gubanc alakot öltenek. Következésképpen a másodlagos szerkezetek stabilitását az interpeptid hidrogénkötések határozzák meg. Más típusú kötések ebben szinte nem vesznek részt, kivéve a diszulfidkötéseket a polipeptidlánc mentén a cisztein-maradékok helyein. A rövid peptidek a diszulfid kötések miatt ciklusokba záródnak. Sok fehérje α-hélikális régiókat és β-struktúrákat egyaránt tartalmaz. Szinte nincs 100%-ban α-hélixből álló természetes fehérje (kivétel a paramiozin, egy 96-100%-ban α-hélixből álló izomfehérje), míg a szintetikus polipeptidek 100%-ban hélixből állnak.
Más fehérjék különböző fokú tekercseléssel rendelkeznek. Az α-helikális struktúrák nagy gyakorisága figyelhető meg a paramiozinban, a mioglobinban és a hemoglobinban. Ezzel szemben a tripszinben, egy ribonukleázban a polipeptidlánc jelentős része réteges β-struktúrákká hajtódik. A támasztószövetek fehérjéi: keratin (szőr, gyapjú fehérje), kollagén (inak, bőr fehérje), fibroin (természetes selyem fehérje) polipeptid láncokból álló β-konfigurációjú. A fehérjék polipeptidláncainak különböző fokú helicitása azt jelzi, hogy nyilvánvalóan vannak olyan erők, amelyek részben megzavarják a helicitást, vagy „megtörik” a polipeptidlánc szabályos feltekeredését. Ennek az az oka, hogy a fehérje polipeptidlánc egy bizonyos térfogatban, azaz egy harmadlagos szerkezetbe tömörödik.
A fehérje harmadlagos szerkezete
A fehérje harmadlagos szerkezete a polipeptidlánc térbeli elrendezésének módja. A fehérjék harmadlagos szerkezetük alakja alapján főként globulárisra és fibrillárisra oszthatók. A globuláris fehérjék leggyakrabban ellipszoid alakúak, a fibrilláris (szálszerű) fehérjék pedig megnyúlt alakúak (rúd vagy orsó alakú).
A fehérjék harmadlagos szerkezetének konfigurációja azonban még nem ad okot arra gondolni, hogy a fibrilláris fehérjéknek csak β-szerkezetük van, a globuláris fehérjéknek pedig α-helikális szerkezetük van. Vannak olyan fibrilláris fehérjék, amelyek spirális, nem pedig rétegzett, hajtogatott másodlagos szerkezettel rendelkeznek. Például az α-keratin és a paramiozin (a puhatestűek elzáró izmának fehérje), a tropomiozinok (a vázizmok fehérjéi) a fibrilláris fehérjékhez tartoznak (rúd alakúak), másodlagos szerkezetük α-hélix; ezzel szemben a globuláris fehérjék nagyszámú β-struktúrát tartalmazhatnak.
Egy lineáris polipeptidlánc spiralizációja körülbelül négyszeresére csökkenti a méretét; a harmadlagos szerkezetbe való bepakolás pedig az eredeti láncnál több tízszer tömörebbé teszi.
A fehérje harmadlagos szerkezetét stabilizáló kötések. Az aminosavak oldalgyökei közötti kötések szerepet játszanak a tercier szerkezet stabilizálásában. Ezek a kapcsolatok a következőkre oszthatók:
erős (kovalens) [előadás].
gyenge (poláris és van der Waals) [előadás].
Az aminosavak oldalgyökei között számos kötés határozza meg a fehérjemolekula térbeli konfigurációját.
A fehérje harmadlagos szerkezetének szerveződésének jellemzői. A polipeptid lánc harmadlagos szerkezetének konformációját a benne található aminosavak oldalgyökeinek tulajdonságai (amelyek nem gyakorolnak észrevehető hatást az elsődleges és másodlagos struktúrák kialakulására) és a mikrokörnyezet, azaz a környezet. Hajlított állapotban a fehérje polipeptidlánca hajlamos energetikailag kedvező formát ölteni, amelyet minimális szabadenergia jellemez. Ezért a nem poláris R-csoportok, „elkerülve” a vizet, mintegy a fehérje harmadlagos szerkezetének belső részét alkotják, ahol a polipeptidlánc hidrofób oldalláncainak fő része található. A fehérjegömb közepén szinte nincs vízmolekula. Az aminosav poláris (hidrofil) R csoportjai ennek a hidrofób magnak a külsején helyezkednek el, és vízmolekulákkal vannak körülvéve. A polipeptidlánc háromdimenziós térben bonyolultan meg van hajlítva. Amikor meghajlik, a másodlagos spirális konformáció megszakad. A lánc azokon a gyenge pontokon „szakad meg”, ahol a prolin vagy a hidroxiprolin található, mivel ezek az aminosavak mozgékonyabbak a láncban, és csak egy hidrogénkötést képeznek más peptidcsoportokkal. Egy másik hajlítási hely a glicin, amely kis R csoporttal (hidrogén) rendelkezik. Ezért az egyéb aminosavak R-csoportjai egymásra halmozva általában elfoglalják a szabad helyet a glicin helyén. Számos aminosav - alanin, leucin, glutamát, hisztidin - hozzájárul a stabil spirális szerkezetek megőrzéséhez a fehérjékben, és mint például a metionin, valin, izoleucin, aszparaginsav elősegíti a β-struktúrák kialakulását. A tercier konfigurációjú fehérjemolekulában α-hélixek (hélikális), β-struktúrák (réteges) és véletlenszerű tekercsek formájában találhatók. Csak a fehérje helyes térbeli elrendezése teszi aktívvá; megsértése a fehérje tulajdonságainak megváltozásához és a biológiai aktivitás elvesztéséhez vezet.
Kvaterner fehérjeszerkezet
Az egy polipeptidláncból álló fehérjéknek csak harmadlagos szerkezetük van. Ezek közé tartozik a mioglobin - az izomszövet fehérje, amely részt vesz az oxigén megkötésében, számos enzim (lizozim, pepszin, tripszin stb.). Egyes fehérjék azonban több polipeptidláncból épülnek fel, amelyek mindegyike harmadlagos szerkezettel rendelkezik. Az ilyen fehérjéknél bevezették a kvaterner szerkezet fogalmát, amely több, harmadlagos szerkezetű polipeptidlánc egyetlen funkcionális fehérjemolekulává történő szerveződését jelenti. Az ilyen kvaterner szerkezetű fehérjét oligomernek, harmadlagos szerkezetű polipeptidláncait protomereknek vagy alegységeknek nevezzük (4. ábra).

A szerveződés kvaterner szintjén a fehérjék megtartják a harmadlagos szerkezet alapvető konfigurációját (globuláris vagy fibrilláris). Például a hemoglobin egy kvaterner szerkezetű fehérje, és négy alegységből áll. Mindegyik alegység globuláris fehérje, és általában a hemoglobinnak is van globuláris konfigurációja. Haj- és gyapjúfehérjék - a keratinok, amelyek harmadlagos szerkezetükben rokonok a fibrilláris fehérjékkel, fibrilláris konformációval és kvaterner szerkezettel rendelkeznek.
A fehérje kvaterner szerkezetének stabilizálása. Minden kvaterner szerkezetű fehérjét különálló makromolekulák formájában izolálnak, amelyek nem bomlanak alegységekre. Az alegységek felületei közötti érintkezés csak az aminosavak poláris csoportjai miatt lehetséges, mivel az egyes polipeptidláncok harmadlagos szerkezetének kialakulása során a nem poláris aminosavak oldalgyökei (amelyek a polipeptidek többségét alkotják) összes proteinogén aminosav) az alegység belsejében rejtőznek. Poláros csoportjaik között számos ionos (só), hidrogén- és esetenként diszulfidkötés képződik, amelyek szilárdan tartják az alegységeket szervezett komplex formájában. A hidrogénkötéseket megbontó vagy a diszulfidhidakat redukáló anyagok használata a protomerek szétesését és a fehérje kvaterner szerkezetének tönkremenetelét okozza. táblázatban Az 1. ábra a fehérjemolekula különböző szerveződési szintjeit stabilizáló kötésekre vonatkozó adatokat foglalja össze [előadás].
Egyes fibrilláris fehérjék szerkezeti szerveződésének jellemzői
A fibrilláris fehérjék szerkezeti felépítése számos jellemzővel rendelkezik a globuláris fehérjékhez képest. Ezek a tulajdonságok a keratin, a fibroin és a kollagén példáján láthatók. A keratinok α- és β-konformációban léteznek. Az α-Keratinok és a fibroin rétegesen hajtogatott másodlagos szerkezetűek, azonban a keratinban a láncok párhuzamosak, a fibroinban pedig antiparallelek (lásd 3. ábra); Ezenkívül a keratin láncok közötti diszulfid kötéseket tartalmaz, míg a fibroin nem rendelkezik ilyenekkel. A diszulfid kötések felszakadása a polipeptid láncok szétválásához vezet a keratinokban. Éppen ellenkezőleg, a keratinokban a maximális számú diszulfidkötés kialakulása oxidálószerek hatására erős térszerkezetet hoz létre. Általánosságban elmondható, hogy a fibrilláris fehérjékben, a globuláris fehérjékkel ellentétben, néha nehéz szigorúan megkülönböztetni a különböző szerveződési szinteket. Ha elfogadjuk (mint egy globuláris fehérjénél), hogy a tercier szerkezetet egy polipeptid lánc térbeli lefektetésével kell kialakítani, a kvaterner szerkezetet pedig több lánc, akkor a fibrilláris fehérjékben már a másodlagos szerkezet kialakulása során több polipeptid lánc vesz részt. . A fibrilláris fehérje tipikus példája a kollagén, amely az emberi szervezetben az egyik legnagyobb mennyiségben előforduló fehérje (az összes fehérje tömegének körülbelül 1/3-a). A nagy szilárdságú és alacsony nyújthatóságú szövetekben található (csontok, inak, bőr, fogak stb.). A kollagénben az aminosavak egyharmada glicin, és körülbelül egynegyede vagy valamivel több prolin vagy hidroxiprolin.

A kollagén izolált polipeptidlánca (elsődleges szerkezete) szaggatott vonalnak tűnik. Körülbelül 1000 aminosavat tartalmaz, molekulatömege körülbelül 10 5 (5. ábra, a, b). A polipeptidlánc a következő összetételű aminosavak ismétlődő triójából (triplett) épül fel: gly-A-B, ahol A és B a glicintől eltérő aminosavak (leggyakrabban prolin és hidroxiprolin). A kollagén polipeptid láncok (vagy α-láncok) a másodlagos és harmadlagos struktúrák kialakulása során (5. ábra, c és d) nem tudnak tipikus, spirális szimmetriájú α-hélixeket létrehozni. A prolin, a hidroxiprolin és a glicin (antihelikális aminosavak) zavarják ezt. Ezért három α-lánc mintegy csavart spirálokat alkot, mint három szál, amely egy henger köré tekeredne. Három spirális α-lánc alkot egy ismétlődő kollagénszerkezetet, amelyet tropokollagénnek neveznek (5d. ábra). A tropokollagén szervezetében a kollagén harmadlagos szerkezete. A lánc mentén rendszeresen váltakozó prolin és hidroxiprolin lapos gyűrűi merevséget adnak neki, csakúgy, mint a tropokollagén α-láncai közötti láncok közötti kötések (ezért a kollagén ellenáll a nyújtásnak). A tropokollagén lényegében a kollagénrostok egyik alegysége. A tropokollagén alegységek berakása a kollagén kvaterner szerkezetébe lépésenként történik (5e. ábra).
A kollagénszerkezetek stabilizálása a láncok közötti hidrogén-, ion- és van der Waals-kötések, valamint kis számú kovalens kötés miatt következik be.
A kollagén α-láncai eltérő kémiai szerkezettel rendelkeznek. Különböző típusú α 1 láncok (I, II, III, IV) és α 2 láncok léteznek. Attól függően, hogy mely α 1 - és α 2 -láncok vesznek részt a tropokollagén háromszálú hélixének kialakításában, a kollagén négy típusát különböztetjük meg:
az első típus - két α 1 (I) és egy α 2 lánc;
a második típus - három α 1 (II) lánc;
harmadik típus - három α 1 (III) lánc;
negyedik típus - három α 1 (IV) lánc.
A leggyakoribb kollagén az első típus: megtalálható a csontszövetben, a bőrben, az inakban; a második típusú kollagén a porcszövetben stb. található. Egy szövettípusban különböző típusú kollagén lehet.
A kollagén struktúrák rendezett aggregációja, merevsége és tehetetlensége biztosítja a kollagénrostok nagy szilárdságát. A kollagén fehérjék szénhidrát komponenseket is tartalmaznak, azaz fehérje-szénhidrát komplexek.
A kollagén egy extracelluláris fehérje, amelyet minden szervben megtalálható kötőszöveti sejtek képeznek. Ezért a kollagén károsodásával (vagy képződésének megzavarásával) a szervek kötőszövetének támogató funkcióinak többszörös megsértése következik be.
Az alfa polipeptid lánc a valin-leucin aminosav kombinációval, a béta polipeptid lánc pedig a valin-hisztidin-leucin kombinációval végződik. Az alfa- és béta-polipeptid láncok a hemoglobin molekulában nem lineárisan helyezkednek el, ez az elsődleges szerkezet. Az intramolekuláris erők megléte miatt a polipeptidláncok a fehérjékre jellemző alfa-hélix hélix (másodlagos szerkezet) formájában csavarodnak. Maga az alfa-hélix hélix minden alfa- és béta-polipeptidlánchoz térben meghajlik, így tojásdad alakú plexusokat képez (harmadlagos szerkezet). A polipeptidláncok alfa-hélix hélixeinek egyes részeit latin betűkkel jelölik A-tól H-ig. Mind a négy harmadlagos görbületű alfa- és béta-polipeptidlánc térben egy bizonyos viszonyban helyezkedik el – ez egy kvaterner szerkezet. Nem valódi kémiai kötések kötik össze őket, hanem intermolekuláris erők.
Kiderült, hogy az emberben a normál hemoglobin három fő típusa van: az embrionális - U, a magzati - F és a felnőtt hemoglobin - A. A HbU (a méh szó kezdőbetűjéről nevezték el) az embrióban 7 és 12 élethet között fordul elő, majd eltűnik, és megjelenik a magzati hemoglobin, amely a harmadik hónap után a magzat fő hemoglobinja. Ezt követően fokozatosan megjelenik a hétköznapi felnőtt hemoglobin, az úgynevezett HbA, az angol „adult” szó kezdőbetűje után. A magzati hemoglobin mennyisége fokozatosan csökken, így a születés pillanatában a hemoglobin 80%-a HbA és csak 20%-a HbF. Születés után a magzat hemoglobinszintje tovább csökken, és 2-3 életévre már csak 1-2%. Ugyanannyi magzati hemoglobin van egy felnőttben. A 2%-ot meghaladó HbF mennyiség kórosnak minősül felnőtt és 3 év feletti gyermekek esetében.
A normál hemoglobintípusokon kívül jelenleg több mint 50 patológiás változata ismert. Először latin betűkkel nevezték el őket. A B betű hiányzik a hemoglobin típusok jelöléséből, mivel eredetileg a HbS-t jelölte.
Hemoglobin (Hb)- a vörösvértestekben jelenlévő kromoprotein, amely részt vesz az oxigén szövetekbe történő szállításában. A felnőttek hemoglobint hemoglobin A-nak (Hb A) nevezik. Molekulatömege körülbelül 65 000 Da. A Hb A molekula kvaterner szerkezetű, és négy alegységet tartalmaz – polipeptid láncokat (ezek α1, α2, β1 és β2 jelöléssel rendelkeznek, amelyek mindegyike hemhez kapcsolódik.
Ne feledje, hogy a hemoglobin egy allosztérikus fehérje, amelynek molekulái reverzibilisen változhatnak egyik konformációból a másikba. Ez megváltoztatja a fehérje ligandumok iránti affinitását. A ligandumhoz legkisebb affinitású konformációt feszült vagy T-konformációnak nevezzük. A ligandumhoz legnagyobb affinitással rendelkező konformációt relaxált vagy R-konformációnak nevezzük.
A hemoglobin molekula R- és T-konformációi dinamikus egyensúlyi állapotban vannak:
Különféle környezeti tényezők elmozdíthatják ezt az egyensúlyt egyik vagy másik irányba. A Hb O2 iránti affinitását befolyásoló alloszterikus szabályozók a következők: 1) oxigén; 2) H+ koncentráció (közepes pH); 3) szén-dioxid (CO2); 4) 2,3-difoszfoglicerát (DPG). Az oxigénmolekulának a hemoglobin egyik alegységéhez való kapcsolódása elősegíti a feszült konformáció átmenetét egy laza konformációba, és növeli ugyanazon hemoglobin molekula más alegységeinek oxigén iránti affinitását. Ezt a jelenséget kooperatív hatásnak nevezzük. A hemoglobin oxigénhez való kötődésének összetett jellegét a hemoglobin O2 telítési görbéje tükrözi, amely S alakú (3.1. ábra).

3.1. ábra. A mioglobin (1) és a hemoglobin (2) oxigéntelítettség görbéi.
Molekuláris formák A hemoglobin a polipeptid láncok szerkezetében különbözik egymástól. A fiziológiás körülmények között létező hemoglobin ilyen sokféleségére példa az magzati hemoglobin (HbF), amelyek jelen vannak a vérben az emberi fejlődés embrionális szakaszában. A HbA-val ellentétben molekulája 2 α- és 2 γ-láncot tartalmaz (vagyis a β-láncokat γ-láncok helyettesítik). Az ilyen hemoglobinnak nagyobb az affinitása az oxigénhez. Ez az, ami lehetővé teszi, hogy az embrió oxigént kapjon az anya véréből a placentán keresztül. Nem sokkal a születés után a csecsemő vérében lévő HbF-et HbA váltja fel.
A kóros vagy patológiás hemoglobinra példa a már említett (lásd 2.4.) hemoglobin S, amely sarlósejtes vérszegénységben szenvedő betegeknél található. Amint azt már tudja, abban különbözik a hemoglobin A-tól, hogy a β-láncokban a glutamátot valinnal helyettesíti. Ez az aminosav-szubsztitúció a HbS vízben való oldhatóságának csökkenését és az O2 iránti affinitásának csökkenését okozza.
Szerves anyagok. Az élő szervezetek a szervetleneken kívül sokféle szerves anyagot is tartalmaznak. Az élőlények szerves anyagát elsősorban négy kémiai elem alkotja, ún biogén: szén, hidrogén, oxigén és nitrogén. A fehérjékben ezekhez az elemekhez ként, a nukleinsavakban pedig foszfort adnak.
A szerves anyagok sokféleségét nagymértékben a szén határozza meg. Egyedülálló tulajdonságainak köszönhetően ez az elem az élet kémiai alapját képezi. Számos atommal és csoportjaival kovalens kötéseket tud kialakítani, láncokat és gyűrűket képezve, amelyek különböző összetételű, szerkezetű, hosszúságú és alakú szerves molekulák vázát alkotják. Ezek viszont összetett kémiai vegyületeket képeznek, amelyek szerkezetükben és funkciójukban különböznek egymástól. A szerves molekulák sokféleségének fő oka nem annyira az őket alkotó atomok különbsége, hanem sokkal inkább a molekulában való elrendeződésük eltérő sorrendje.
A biopolimerek fogalma. Egy élő szervezetben a szerves anyagokat vagy kis molekulák, viszonylag alacsony molekulatömegűek, vagy nagy makromolekulák képviselik. Az alacsony molekulatömegű vegyületek közé tartoznak az aminosavak, cukrok, szerves savak, alkoholok, vitaminok stb.
A fehérjék, poliszacharidok és nukleinsavak többnyire nagy molekulatömegű szerkezetek. Ezért hívják őket makrómolekulák(görögből makró- nagy). Így a legtöbb fehérje molekulatömege 5 000 és 1 000 000 közötti nagy molekulatömegű szerves vegyületeket - fehérjéket, nukleinsavakat, poliszacharidokat, amelyek molekulái nagyszámú, azonos vagy eltérő kémiai szerkezetű ismétlődő egységből állnak. biopolimerek(görögből bios- élet és irányelv– számos). Azokat az egyszerű molekulákat, amelyekből a biopolimerek épülnek, nevezzük monomerek. A fehérjék monomerei aminosavak, a poliszacharidok monomerei monoszacharidok, a nukleinsavak monomerei pedig nukleotidok. A makromolekulák a sejt száraz tömegének körülbelül 90%-át teszik ki.
Ez a fejezet a makromolekulák mindhárom osztályát és azok monomer egységeit vizsgálja. A lipideket is figyelembe vették - molekulák, amelyek általában sokkal kisebbek, mint a biopolimerek, de funkciókat is ellátnak a szervezetben.
A szerves anyagok egy speciális csoportja biológiailag aktív anyagok: enzimek, hormonok, vitaminok stb. Szerkezetükben változatosak; befolyásolja az anyagcserét és az energiaátalakítást.
A különböző szervezetcsoportok sejtjeiben bizonyos szerves vegyületek tartalma eltérő. Például az állati sejtekben a fehérjék és zsírok, míg a növényi sejtekben a szénhidrátok. Különböző sejtekben azonban bizonyos szerves vegyületek hasonló funkciókat látnak el.
Mókusok. Az élő szervezetekben funkcionális jelentőségüket tekintve a fehérjék játsszák a vezető szerepet a makromolekulák között. A fehérjék dominánsak és mennyiségileg sok szervezetben vannak. Így az állatok testében száraz tömegük 40-50%-át, a növények testében - 20-35%-át teszik ki. A fehérjék olyan heteropolimerek, amelyek monomerjei aminosavak.
Az aminosavak a fehérjemolekulák „építőkövei”. Aminosavak - bázikus tulajdonságokkal jellemezhető aminocsoportot (–NH) és savas tulajdonságokkal rendelkező karboxilcsoportot (–COOH) egyaránt tartalmazó szerves vegyületek. Az aminocsoport és a karboxilcsoport ugyanahhoz a szénatomhoz kapcsolódik (ábra). E tulajdonság szerint minden aminosav hasonló egymáshoz. A legtöbb fehérjeképző aminosavnak egy karboxilcsoportja és egy aminocsoportja van; ezeket az aminosavakat nevezik semleges.
A molekula része, az úgynevezett radikális (R) a különböző aminosavak eltérő szerkezetűek (ábra). A különböző aminosavak gyökök lehetnek nempolárisak vagy polárisak (töltött vagy töltetlenek), hidrofóbok vagy hidrofilek, ami bizonyos tulajdonságokat ad a fehérjéknek. A semlegesek mellett vannak bázikus aminosavak- egynél több aminocsoporttal, valamint savas aminosavak- egynél több karboxilcsoporttal. Egy további amino- vagy hidroxilcsoport jelenléte befolyásolja a gyök tulajdonságait. Az aminosavgyökök minden tulajdonsága döntő szerepet játszik a fehérje térszerkezetének kialakításában.
Az összes ismert aminosav száma körülbelül 200, és csak 20 típus vesz részt a természetes fehérjék képzésében. Ezeket az aminosavakat ún fehérjeképző(2. táblázat; a táblázat az aminosavak teljes és rövidített nevét mutatja, nem memorizálás céljából).
2. táblázat: Bázikus aminosavak és rövidítéseik
A növények és baktériumok a fotoszintézis elsődleges termékeiből minden szükséges aminosavat szintetizálhatnak. Az ember és az állat nem képes minden aminosavat szintetizálni, ezért az ún esszenciális aminosavakétellel készen kell fogadni.
Az ember számára esszenciális aminosavak a következők: lizin, valin, leucin, izoleucin, treonin, fenilalanin, triptofánÉs metionin; gyermekek számára is nélkülözhetetlenek argininÉs hisztidin. Az összes esszenciális aminosavat tartalmazó élelmiszer-fehérjéket ún teljes értékű, Nem úgy mint fogyatékos, amelyekből hiányoznak bizonyos esszenciális aminosavak.
A bázikus és savas csoportok jelenléte egy aminosavban meghatározza azok amfoterségét és nagy reakcióképességét. Amino csoport
Az egyik aminosav (–NH 2) kölcsönhatásba léphet egy másik aminosav karboxilcsoportjával (–COOH), vízmolekulát szabadítva fel. A kapott molekula az dipeptid (ábra), és a –CO-NH– kötést ún peptid. A dipeptidmolekula egyik végén egy szabad aminocsoport, a másik végén egy karboxilcsoport található. Ennek köszönhetően a dipeptid más aminosavakat is tud magához kötni, kialakítva oligopeptidek. Ha sok aminosav kapcsolódik ilyen módon (több mint tíz), akkor hosszú lánc jön létre - polipeptid.
A peptidek fontos szerepet játszanak a szervezetben. Sok oligo- és polipeptid hormon, antibiotikum és toxin.
Az oligopeptidek közé tartoznak például az oxitocin és a vazopresszin agyalapi mirigyhormonok, valamint a bradikinin (fájdalompeptid) és egyes opiátok (emberi „természetes gyógyszerek”), amelyek fájdalomcsillapítóként hatnak. Rendszeres használat A kábítószer-használat nagyon veszélyes, tönkreteszi a szervezet opiátrendszerét, ezért egy adag kábítószer nélkül szenvedő szenvedélybeteg erős fájdalmat – „elvonást” él át. Az oligopeptidek közé tartozik néhány antibiotikum, például a gramicidin S.
A hormonok (inzulin, adrenokortikotrop hormon stb.), az antibiotikumok (gramicidin A), a toxinok (diphtheria toxin) szintén polipeptidek.
A polipeptidláncok nagyon hosszúak lehetnek, és sokféle aminosav-kombinációt tartalmazhatnak. Azokat a polipeptideket, amelyek molekulája 50-től több ezerig terjedő, 6000 feletti molekulatömegű aminosavmaradékot tartalmaz, fehérjéknek nevezzük.
Minden egyes fehérjét szigorúan állandó összetétel és aminosavsorrend jellemez.
Egy fehérjemolekula szerveződési szintjei. A fehérjemolekulák különböző térbeli formákat ölthetnek – konformáció, amelyek szervezetük négy szintjét képviselik (ábra).
Sok aminosavból álló lánc, amelyet peptidkötések kapcsolnak össze elsődleges szerkezete fehérje molekula. Ez a legfontosabb szerkezet, mivel meghatározza formáját, tulajdonságait és funkcióit. Az elsődleges struktúra alapján más típusú struktúrák jönnek létre. Ezt a szerkezetet kódolja a DNS-molekula. A szervezetben minden egyes fehérje egyedi elsődleges szerkezettel rendelkezik. Egy adott fehérje (például albumin) minden molekulájában ugyanaz az aminosav-csoportok váltakozása, ami megkülönbözteti az albumint bármely más egyedi fehérjétől. Az elsődleges szerkezet sokféleségét a polipeptidláncban lévő aminosavak összetétele, mennyisége és sorrendje határozza meg.
Másodlagos szerkezet A fehérjék a polipeptidlánc különböző aminosavcsoportjainak NH-csoportjainak hidrogénatomja és CO-csoportjainak oxigénatomja közötti hidrogénkötések kialakulásának eredményeként jönnek létre. A polipeptid lánc spirálba van csavarva. A hidrogénkötések ugyan gyengék, de jelentős számuk miatt biztosítják ennek a szerkezetnek a stabilitását. A keratin fehérje molekulák teljesen spirális konfigurációval rendelkeznek. Ez a haj, szőr, karmok, toll és szarv szerkezeti fehérje; a gerincesek bőrének külső rétegének része. A keratin mellett a spirális másodlagos szerkezet a fibrilláris (szálszerű) fehérjékre jellemző, mint például a miozin, a fibrinogén és a kollagén.
A fehérje másodlagos szerkezetét a hélixen kívül egy hajtogatott réteg is ábrázolhatja. Az összehajtott rétegben több polipeptidlánc (vagy egy polipeptidlánc szakasza) párhuzamosan fekszik, harmonikaszerűen hajtogatott lapos konfigurációt alkotva (b6. ábra). Például a fibroin fehérje, amely a selyemhernyó selyemkiválasztó mirigyei által a gubók szövése során kiválasztott selyemrost nagy részét alkotja, másodlagos szerkezetű, hajtogatott réteg formájában.
Harmadlagos szerkezet a cisztein (kéntartalmú aminosav) közötti S-S kötések („diszulfidhidak”), valamint hidrogén-, ionos és egyéb kölcsönhatások révén jön létre. A harmadlagos szerkezet meghatározza a fehérjemolekulák specificitását és biológiai aktivitását. Az olyan fehérjéknek, mint a mioglobin (az izmokban található fehérje; részt vesz az oxigéntartalékok létrehozásában), a tripszin (egy enzim, amely lebontja a belekben a fehérjéket) harmadlagos szerkezetű.
Egyes esetekben több, harmadlagos szerkezetű polipeptidláncot egyetlen komplexté egyesítenek, ezáltal képződnek kvaterner szerkezet. Ebben a fehérje alegységei nincsenek kovalensen kötve, az erősséget gyenge intermolekuláris erők kölcsönhatása biztosítja. Például a kvaterner szerkezet a hemoglobin fehérjére jellemző, amely négy fehérje alegységből és egy nem fehérje részből - a hemből áll.
s 1. Mik azok a fehérjék? 2. Mi a fehérjék szerkezete? 3. Mik azok az aminosavak? 4. Hogyan kapcsolódnak össze az aminosavak, hogy polipeptidláncot képezzenek? 5. Milyen szintjei vannak a fehérjék szerkezeti szerveződésének? 6. Milyen kémiai kötések határozzák meg a fehérjemolekulák szerkezeti szerveződésének különböző szintjeit? 7. Háromféle aminosav létezik A.B.C. Hány öt aminosavból álló polipeptid lánc variáns állítható elő? A polipeptidek ugyanazokkal a tulajdonságokkal rendelkeznek?
Rövid leírás:
Tankönyvtöredék: Biológiai kémia gyakorlatokkal és feladatokkal: tankönyv / szerk. Levelező tag RAMS S.E. Severina. M.: GEOTAR-Média, 2011. - 624 p.: ill. 1. MODUL: A FEHÉRJÉK SZERKEZETE, TULAJDONSÁGAI ÉS FUNKCIÓI
1. MODUL: A FEHÉRJÉK SZERKEZETE, TULAJDONSÁGAI ÉS FUNKCIÓI
Modul felépítés | Témák |
Moduláris egység 1 | 1.1. A fehérjék szerkezeti szerveződése. A natív fehérje konformáció kialakulásának szakaszai 1.2. A fehérje működésének alapjai. Gyógyszerek, mint ligandumok, amelyek befolyásolják a fehérje működését 1.3. A fehérjék denaturációja és spontán renativációjuk lehetősége |
Moduláris egység 2 | 1.4. Az oligomer fehérjék szerkezetének és működésének jellemzői a hemoglobin példáján 1.5. A natív fehérje konformáció fenntartása sejtes körülmények között 1.6. Változatos fehérjék. Fehérjecsaládok az immunglobulinok példájával 1.7. A fehérjék fizikai-kémiai tulajdonságai és elválasztásuk módszerei |
1. moduláris egység A MONOMER FEHÉRJEK SZERKEZETI RENDEZÉSE ÉS MŰKÖDÉSÜK ALAPJAI
Tanulási célok Legyen képes:
1. Használja fel a fehérjék szerkezeti sajátosságaira és a fehérjefunkciók szerkezetétől való függőségére vonatkozó ismereteket az örökletes és szerzett proteinopátiák kialakulásának mechanizmusainak megértéséhez.
2. Ismertesse egyes gyógyszerek terápiás hatásának mechanizmusait, mint ligandumokat, amelyek kölcsönhatásba lépnek a fehérjékkel és megváltoztatják azok aktivitását.
3. Használja a fehérjék szerkezetére és konformációs labilitására vonatkozó ismereteket, hogy megértse szerkezeti és funkcionális instabilitását és denaturálódását változó körülmények között.
4. Ismertesse a denaturáló szerek használatát orvosi anyagok és műszerek sterilizálására, valamint antiszeptikumként.
Tud:
1. A fehérjék szerkezeti szerveződésének szintjei.
2. A fehérjék elsődleges szerkezetének jelentősége, amely meghatározza azok szerkezeti és funkcionális sokféleségét.
3. Az aktív centrum kialakulásának mechanizmusa a fehérjékben és specifikus kölcsönhatása a ligandumokkal, amely a fehérjék működésének hátterében áll.
4. Példák exogén ligandumok (gyógyszerek, toxinok, mérgek) hatására a fehérjék konformációjára és funkcionális aktivitására.
5. A fehérje denaturáció okai és következményei, denaturációt okozó tényezők.
6. Példák a denaturáló faktorok alkalmazására az orvostudományban antiszeptikumként és orvosi műszerek sterilizálására.
TÉMAKÖR 1.1. A FEHÉRJÉK SZERKEZETI RENDEZÉSE. AZ ÖSSZEFÜGGÉS KIALAKULÁSÁNAK SZAKASZAI
FEHÉRJE KONFORMÁCIÓK
Mókusok - ezek heteropolimerek molekulák (azaz sokféle monomerből áll). A fehérjemonomerek 20 típusú α-aminosav, amelyek peptidkötésekkel kapcsolódnak egymáshoz.
A fehérjében lévő aminosavak kombinációját és sorrendjét az egyedek DNS-ében lévő megfelelő gének szerkezete határozza meg. Minden fehérje sajátos szerkezetének megfelelően ellátja saját funkcióját. Egy adott szervezet fehérjéinek halmaza ( proteom) meghatározza annak fenotípusos jellemzőit, valamint az örökletes betegségek jelenlétét vagy azok kialakulására való hajlamot.
1. A fehérjéket alkotó aminosavak. Peptid kötés.
A fehérjék monomerekből - 20 α-aminosavból - felépített heteropolimerek.
Az aminosavak általános képlete az alábbiakban látható.

Az aminosavak szerkezetükben, méretükben és az α-szénatomhoz kapcsolódó gyökök fizikai-kémiai tulajdonságaiban különböznek egymástól. Az aminosavak funkciós csoportjai meghatározzák a különböző α-aminosavak tulajdonságainak jellemzőit. Az α-aminosavakban található gyökök több csoportra oszthatók:
prolin, A többi 19 fehérjemonomertől eltérően nem aminosav, hanem egy iminosav, a prolinban lévő gyök mind az α-szénatomhoz, mind az iminocsoporthoz kapcsolódik

Az aminosavak vízben való oldhatósága változó. Ez annak köszönhető, hogy a gyökök képesek kölcsönhatásba lépni a vízzel (hidráttal).
NAK NEK hidrofil Ide tartoznak az anionos, kationos és poláris töltés nélküli funkciós csoportokat tartalmazó gyökök.
NAK NEK hidrofób metilcsoportokat, alifás láncokat vagy gyűrűket tartalmazó gyököket foglalnak magukban.
2. A peptidkötések aminosavakat kapcsolnak össze peptidekké. A peptidszintézis során az egyik aminosav α-karboxilcsoportja kölcsönhatásba lép egy másik aminosav α-aminocsoportjával, így képződik peptid kötés:
A fehérjék polipeptidek, azaz. α-aminosavak peptidkötéssel összekapcsolt lineáris polimerjei (1.1. ábra)
 Rizs. 1.1. A peptidek szerkezetének leírására használt kifejezések
Rizs. 1.1. A peptidek szerkezetének leírására használt kifejezések
A polipeptideket alkotó aminosavak monomereit ún aminosav-maradékok. Ismétlődő csoportok lánca - NH-CH-CO- nyomtatványok peptid gerinc. A szabad α-aminocsoporttal rendelkező aminosavakat N-terminálisnak, a szabad α-karboxilcsoportot C-terminálisnak nevezzük. A peptideket az N-terminálistól a C-terminálisig írják és olvassák.
A prolin iminocsoportja által létrehozott peptidkötés eltér a többi peptidkötéstől: a peptidcsoport nitrogénatomjából hiányzik a hidrogén,
ehelyett egy gyökös kötés jön létre, aminek következtében a gyűrű egyik oldala a peptidvázba kerül:

A peptidek különböznek az aminosav-összetételben, az aminosavak számában és az aminosavak kapcsolódási sorrendjében, például a Ser-Ala-Glu-Gis és a His-Glu-Ala-Ser két különböző peptid.
A peptidkötések nagyon erősek, kémiai nem enzimatikus hidrolízisük kemény körülményeket igényel: az elemzett fehérjét tömény sósavban hidrolizálják körülbelül 110 °C hőmérsékleten 24 órán át. Egy élő sejtben a peptidkötések megszakadhatnak proteolitikus enzimek, hívott proteázok vagy peptid hidrolázok.
3. A fehérjék elsődleges szerkezete. A különböző fehérjék peptidláncában az aminosavak nem véletlenszerűen váltakoznak, hanem meghatározott sorrendben helyezkednek el. A polipeptid lánc aminosavainak lineáris szekvenciáját vagy váltakozási sorrendjét nevezzük a fehérje elsődleges szerkezete.
Az egyes fehérjék elsődleges szerkezetét a DNS-molekula kódolja (a génnek nevezett régióban), és a transzkripció (az információ mRNS-re másolása) és a transzláció (a fehérje elsődleges szerkezetének szintézise) során valósul meg. Ebből következően az egyén fehérjéinek elsődleges szerkezete a szülőktől a gyerekeknek örökletesen átadott információ, amely meghatározza az adott szervezet fehérjéinek szerkezeti sajátosságait, amelyektől a meglévő fehérjék működése függ (1.2. ábra).
 Rizs. 1.2. A genotípus és az egyén szervezetében szintetizált fehérjék konformációja közötti kapcsolat
Rizs. 1.2. A genotípus és az egyén szervezetében szintetizált fehérjék konformációja közötti kapcsolat
Az emberi szervezetben található körülbelül 100 000 egyedi fehérje mindegyike rendelkezik egyedi elsődleges szerkezete. Az azonos típusú fehérje (például albumin) molekuláiban az aminosavak azonos váltakozása található, ami megkülönbözteti az albumint bármely más egyedi fehérjétől.
A peptidláncban lévő aminosavak szekvenciája az információrögzítés egy formájának tekinthető. Ez az információ határozza meg a lineáris peptidlánc térbeli elrendeződését egy kompaktabb háromdimenziós szerkezetté, ún. konformáció mókus. A funkcionálisan aktív fehérje konformáció kialakulásának folyamatát ún összecsukható
4. Fehérje konformáció. A peptidvázban szabad forgás lehetséges a peptidcsoport nitrogénatomja és a szomszédos α-szénatom, valamint az α-szénatom és a karbonilcsoport szénatomja között. Az aminosavmaradékok funkcionális csoportjainak kölcsönhatása miatt a fehérjék primer szerkezete bonyolultabb térszerkezetekre tehet szert. A globuláris fehérjékben a peptidláncok konformációjának két fő szintje van: másodlagosÉs harmadlagos szerkezet.
A fehérjék másodlagos szerkezete a peptidváz -C=O és -NH- funkciós csoportjai közötti hidrogénkötések létrejöttének eredményeként kialakuló térszerkezet. Ebben az esetben a peptidlánc kétféle szabályos szerkezetet szerezhet: α-hélixekÉs β-struktúrák.
BAN BEN α-hélixek hidrogénkötések jönnek létre a karbonilcsoport oxigénatomja és a belőle származó 4. aminosav amid-nitrogénjének hidrogénje között; aminosavak oldalláncai
a spirál perifériáján helyezkednek el, anélkül, hogy részt vennének a másodlagos szerkezet kialakításában (1.3. ábra).
Az ömlesztett gyökök vagy azonos töltésű gyökök megakadályozzák az α-hélix kialakulását. A gyűrűs szerkezetű prolin megszakítja az α-hélixet, mivel a peptidláncban a nitrogénatomnál hiányzó hidrogén miatt lehetetlen hidrogénkötést kialakítani. A nitrogén és az α-szénatom közötti kötés a prolingyűrű része, így a peptid gerince ezen a ponton meghajlik.
β-Struktúra Az egyik polipeptid lánc peptidvázának lineáris régiói között képződik, és ezáltal hajtogatott struktúrákat hoz létre. Polipeptidláncok vagy azok részei képződhetnek párhuzamos vagy antiparallel β-struktúrák. Az első esetben a kölcsönhatásba lépő peptidláncok N- és C-terminálisai egybeesnek, a másodikban pedig ellenkező irányúak (1.4. ábra).
Rizs. 1.3. A fehérje másodlagos szerkezete - α-hélix
 Rizs. 1.4. Párhuzamos és antiparallel β-lemez szerkezetek
Rizs. 1.4. Párhuzamos és antiparallel β-lemez szerkezetek
A β-struktúrákat széles nyilak jelzik: A - Antiparallel β-struktúra. B - Párhuzamos β-lemez szerkezetek
Egyes fehérjékben β-struktúrák alakulhatnak ki a különböző polipeptidláncok peptidvázának atomjai közötti hidrogénkötések kialakulásának köszönhetően.
A fehérjékben is megtalálható szabálytalan másodlagos területek szerkezet, amely magában foglalja a polipeptid gerincének hajlításait, hurkjait és fordulatait. Gyakran olyan helyeken helyezkednek el, ahol a peptidlánc iránya megváltozik, például ha párhuzamos β-lemez szerkezet alakul ki.
Az α-hélixek és β-struktúrák jelenléte alapján a globuláris fehérjék négy kategóriába sorolhatók.
Rizs. 1.5. A mioglobin (A) és a hemoglobin β-lánc (B) másodlagos szerkezete, amely nyolc α-hélixet tartalmaz

 Rizs. 1.6. A trioszfoszfát izomeráz és a piruvát kináz domén másodlagos szerkezete
Rizs. 1.6. A trioszfoszfát izomeráz és a piruvát kináz domén másodlagos szerkezete
 Rizs. 1.7. Az immunglobulin (A) konstans doménjének másodlagos szerkezete és a szuperoxid-diszmutáz enzim (B)
Rizs. 1.7. Az immunglobulin (A) konstans doménjének másodlagos szerkezete és a szuperoxid-diszmutáz enzim (B)
BAN BEN negyedik kategória olyan fehérjéket tartalmaztak, amelyek kis mennyiségű szabályos másodlagos struktúrát tartalmaznak. Ezek a fehérjék közé tartoznak a kis ciszteinben gazdag fehérjék vagy metalloproteinek.
A fehérje harmadlagos szerkezete- aminosavgyökök közötti kölcsönhatások következtében kialakuló konformáció, amely a peptidláncban egymástól jelentős távolságra is elhelyezkedhet. A legtöbb fehérje gömbölyűre emlékeztető térszerkezetet alkot (globuláris fehérjék).
Mivel a hidrofób aminosav gyökök hajlamosak egyesülni az ún hidrofób kölcsönhatásokés intermolekuláris van der Waals erők hatására a fehérjegömb belsejében sűrű hidrofób mag képződik. A hidrofil ionizált és nem ionizált gyökök elsősorban a fehérje felszínén helyezkednek el, és meghatározzák annak vízoldhatóságát.
 Rizs. 1.8. A fehérje harmadlagos szerkezetének kialakulása során az aminosavgyökök között létrejövő kötések típusai
Rizs. 1.8. A fehérje harmadlagos szerkezetének kialakulása során az aminosavgyökök között létrejövő kötések típusai
1 - ionos kötés- pozitív és negatív töltésű funkciós csoportok között fordul elő;
2 - hidrogén kötés- hidrofil töltés nélküli csoport és bármely más hidrofil csoport között fordul elő;
3 - hidrofób kölcsönhatások- hidrofób gyökök között keletkeznek;
4 - diszulfid kötés- a cisztein-maradékok SH-csoportjainak oxidációja és egymással való kölcsönhatása következtében jön létre
A hidrofób mag belsejében található hidrofil aminosavak kölcsönhatásba léphetnek egymással iónÉs hidrogénkötések(1.8. ábra).
Az ionos és hidrogénkötések, valamint a hidrofób kölcsönhatások gyengék: energiájuk nem sokkal nagyobb, mint a molekulák szobahőmérsékleten történő hőmozgásának energiája. A fehérje konformációját sok ilyen gyenge kötés kialakítása tartja fenn. Mivel a fehérjét alkotó atomok állandó mozgásban vannak, előfordulhat, hogy egyes gyenge kötések megszakadnak, mások pedig kialakulhatnak, ami a polipeptidlánc egyes szakaszainak enyhe elmozdulásához vezet. A fehérjéknek ezt a tulajdonságát, hogy egyesek felszakadása és más gyenge kötések kialakulása következtében konformációt változtatnak, az ún. konformációs labilitás.
Az emberi testnek vannak olyan rendszerei, amelyek támogatják homeosztázis- a belső környezet állandósága bizonyos, az egészséges szervezet számára elfogadható határokon belül. Homeosztázis körülmények között a konformáció kis változásai nem zavarják meg a fehérjék általános szerkezetét és működését. A fehérje funkcionálisan aktív konformációját ún natív felépítés. A belső környezet változásai (például a glükóz, Ca-ionok, protonok stb. koncentrációja) a konformáció megváltozásához és a fehérjefunkciók megzavarásához vezetnek.
Egyes fehérjék harmadlagos szerkezete stabilizálódik diszulfid kötések, két aminosavból álló -SH csoportok kölcsönhatása következtében jött létre
 Rizs. 1.9. Diszulfid kötés kialakulása fehérjemolekulában
Rizs. 1.9. Diszulfid kötés kialakulása fehérjemolekulában
cisztein (1.9. ábra). A legtöbb intracelluláris fehérje tercier szerkezetében nincs kovalens diszulfidkötés. Jelenlétük a sejt által kiválasztott fehérjékre jellemző, ami biztosítja azok nagyobb stabilitását extracelluláris körülmények között. Így diszulfid kötések vannak jelen az inzulin és az immunglobulinok molekuláiban.
Inzulin- a hasnyálmirigy β-sejtjeiben szintetizált fehérjehormon, amely a vér glükózkoncentrációjának növekedésére válaszul választódik ki a vérben. Az inzulin szerkezetében két diszulfid kötés köti össze a polipeptid A és B láncát, és egy diszulfid kötés az A láncon belül (1.10. ábra).
 Rizs. 1.10. Diszulfid kötések az inzulin szerkezetében
Rizs. 1.10. Diszulfid kötések az inzulin szerkezetében
5. A fehérjék szuperszekunder szerkezete. Az eltérő elsődleges szerkezetű és funkciójú fehérjékben néha kimutathatók a másodlagos struktúrák hasonló kombinációi és egymáshoz viszonyított helyzetei, amelyeket szuperszekunder szerkezetnek nevezünk. Közbenső helyet foglal el a másodlagos és harmadlagos szerkezetek között, mivel a másodlagos szerkezet elemeinek specifikus kombinációja a fehérje harmadlagos szerkezetének kialakításában. A szuperszekunder struktúráknak sajátos neveik vannak, például „α-helix-turn-a-helix”, „leucin-cipzár”, „cink ujjak” stb. Az ilyen szuperszekunder struktúrák a DNS-kötő fehérjékre jellemzőek.
– Leucin cipzár. Ez a fajta szuperszekunder szerkezet két fehérje összekapcsolására szolgál. A kölcsönhatásba lépő fehérjék felületén α-helikális régiók találhatók, amelyek legalább négy leucinmaradékot tartalmaznak. Az α-hélixben lévő leucinmaradékok hat aminosavra helyezkednek el egymástól. Mivel az α-hélix minden fordulója 3,6 aminosavból áll, ezért minden második körben leucin gyökök találhatók. Egy fehérje α-hélixének leucin-maradékai kölcsönhatásba léphetnek egy másik fehérje leucin-maradékaival (hidrofób kölcsönhatások), összekapcsolva azokat (1.11. ábra). Sok DNS-kötő fehérje oligomer komplexekben működik, ahol az egyes alegységek „leucin cipzárral” kapcsolódnak egymáshoz.
 Rizs. 1.11. "Leucin cipzár" két fehérje α-helikális régiói között
Rizs. 1.11. "Leucin cipzár" két fehérje α-helikális régiói között
Ilyen fehérjék például a hisztonok. Hisztonok- nukleáris fehérjék, amelyek nagyszámú pozitív töltésű aminosavat tartalmaznak - arginint és lizint (akár 80%). A hiszton molekulákat nyolc monomert tartalmazó oligomer komplexekké egyesítik „leucin cipzárak” segítségével, annak ellenére, hogy ezek a molekulák jelentős homonim töltéssel rendelkeznek.
"Cink ujj"- a DNS-kötő fehérjékre jellemző szuperszekunder szerkezetű variáns, a fehérje felületén megnyúlt fragmentum formájában van, és körülbelül 20 aminosavból áll (1.12. ábra). A „kiterjesztett ujj” formát egy cinkatom támogatja, amely négy aminosavcsoporthoz – két cisztein- és két hisztidin-csoporthoz – kötődik. Egyes esetekben a hisztidin-maradékok helyett cisztein-maradékok vannak. Két szorosan fekvő cisztein-maradékot egy körülbelül 12 aminosavból álló Cys-szekvencia választ el a másik két Gisili-maradéktól. A fehérjének ez a régiója α-hélixet alkot, melynek gyökei specifikusan kötődhetnek a DNS fő barázdájának szabályozó régióihoz. Egyedi kötődési specifitás
 Rizs. 1.12. A „cink ujj” szerkezetet alkotó DNS-kötő fehérjék régiójának elsődleges szerkezete (a betűk a szerkezetet alkotó aminosavakat jelölik)
Rizs. 1.12. A „cink ujj” szerkezetet alkotó DNS-kötő fehérjék régiójának elsődleges szerkezete (a betűk a szerkezetet alkotó aminosavakat jelölik)
A szabályozó DNS-kötő fehérje a cinkujj régióban található aminosavak szekvenciájától függ. Az ilyen struktúrák különösen a transzkripció szabályozásában részt vevő szteroid hormonok receptorait tartalmazzák (DNS-ből RNS-be olvasva információkat).
TÉMAKÖR 1.2. A FEHÉRJÉNEK MŰKÖDÉSÉNEK ALAPJAI. GYÓGYSZEREK MINT LIGANDUMOK A FEHÉRJE FUNKCIÓT BEFOLYÁSOLÓ GYÓGYSZEREK
1. A fehérje aktív centruma és kölcsönhatása a ligandummal. A harmadlagos szerkezet kialakulása során egy funkcionálisan aktív fehérje felületén, általában egy mélyedésben, régió jön létre, amelyet az elsődleges szerkezetben egymástól távol eső aminosavgyökök alkotnak. Ezt a régiót, amelynek egyedi szerkezete van egy adott fehérje számára, és képes specifikusan kölcsönhatásba lépni egy adott molekulával vagy hasonló molekulák csoportjával, fehérje-ligandum kötőhelynek vagy aktív helynek nevezzük. A ligandumok olyan molekulák, amelyek kölcsönhatásba lépnek a fehérjékkel.
Magas specifitás A fehérje és a ligand kölcsönhatását az aktív centrum szerkezetének a ligandum szerkezetével való komplementaritása biztosítja.
Komplementaritás- ez a kölcsönható felületek térbeli és kémiai megfeleltetése. Az aktív centrumnak nemcsak térben kell megfelelnie a benne lévő ligandumnak, hanem kötéseknek (ionos, hidrogén- és hidrofób kölcsönhatások) is létre kell jönniük az aktív centrumban lévő gyökök funkciós csoportjai és a ligandumot tartó ligandum között. az aktív központban (1.13. ábra).
 Rizs. 1.13. A fehérje és a ligand komplementer kölcsönhatása
Rizs. 1.13. A fehérje és a ligand komplementer kölcsönhatása
Egyes ligandumok, ha egy fehérje aktív központjához kapcsolódnak, kisegítő szerepet játszanak a fehérjék működésében. Az ilyen ligandumokat kofaktoroknak, a nem fehérje részt tartalmazó fehérjéket pedig nevezzük komplex fehérjék(szemben az egyszerű fehérjékkel, amelyek csak a fehérje részből állnak). A fehérjéhez szorosan kapcsolódó, nem fehérje részt ún protézis csoport. Például a mioglobin, a hemoglobin és a citokrómok tartalmaznak egy vasiont tartalmazó protéziscsoportot, a hemet, amely szilárdan az aktív központhoz kapcsolódik. A hem tartalmú komplex fehérjéket hemoproteineknek nevezzük.
Amikor specifikus ligandumok kapcsolódnak a fehérjékhez, ezeknek a fehérjéknek a funkciója megnyilvánul. Így az albumin, a vérplazma legfontosabb fehérje, úgy fejti ki transzport funkcióját, hogy az aktív centrumhoz hidrofób ligandumokat, például zsírsavakat, bilirubint, egyes gyógyszereket stb. köt (1.14. ábra).
A peptidlánc háromdimenziós szerkezetével kölcsönhatásba lépő ligandumok nem csak kis molekulatömegű szerves és szervetlen molekulák, hanem makromolekulák is lehetnek:
DNS (példák a fentebb tárgyalt DNS-kötő fehérjékre);
poliszacharidok;
 Rizs. 1.14. A genotípus és a fenotípus kapcsolata
Rizs. 1.14. A genotípus és a fenotípus kapcsolata
Az emberi fehérjék egyedi elsődleges szerkezete, amelyet a DNS-molekula kódol, a sejtekben egyedi konformáció, aktív centrum szerkezet és fehérjefunkciók formájában valósul meg.
Ezekben az esetekben a fehérje felismeri a ligandum egy specifikus régióját, amely arányos és komplementer a kötőhellyel. Így a hepatociták felszínén az inzulin hormon receptorfehérjék találhatók, amely fehérjeszerkezettel is rendelkezik. Az inzulin és a receptor kölcsönhatása változást okoz annak konformációjában és aktiválja a jelátviteli rendszereket, ami a tápanyagok tárolását eredményezi a májsejtekben étkezés után.
És így, A fehérjék működése a fehérje aktív centrumának a ligandummal való specifikus kölcsönhatásán alapul.
2. A domén szerkezete és szerepe a fehérjék működésében. A globuláris fehérjék hosszú polipeptidláncai gyakran több kompakt, viszonylag független régióba hajtódnak össze. Független harmadlagos szerkezettel rendelkeznek, amely a globuláris fehérjékre emlékeztet, és ún domainek. A fehérjék doménszerkezetének köszönhetően könnyebben kialakul harmadlagos szerkezetük.
A doménfehérjékben a ligandumkötő helyek gyakran a domének között helyezkednek el. Így a tripszin egy proteolitikus enzim, amelyet a hasnyálmirigy exokrin része termel, és szükséges az élelmiszer-fehérjék emésztéséhez. Kétdoménes szerkezetű, és a tripszin ligandumához - élelmiszerfehérjéhez - való kötődési központja a két domén közötti barázdában található. Az aktív központban létrejönnek a szükséges feltételek az élelmiszer-fehérje egy adott helyének hatékony kötődéséhez és peptidkötéseinek hidrolíziséhez.
A fehérjében lévő különböző domének egymáshoz képest elmozdulhatnak, amikor az aktív centrum kölcsönhatásba lép a ligandummal (1.15. ábra).
Hexokináz- egy enzim, amely ATP segítségével katalizálja a glükóz foszforilációját. Az enzim aktív helye a két domén közötti hasadékban található. Amikor a hexokináz glükózhoz kötődik, a körülötte lévő domének bezáródnak, és a szubsztrát beszorul, ahol foszforiláció történik (lásd 1.15. ábra).
 Rizs. 1.15. A hexokináz domének kötődése glükózhoz
Rizs. 1.15. A hexokináz domének kötődése glükózhoz
Egyes fehérjékben a domének független funkciókat látnak el, különféle ligandumokhoz kötődve. Az ilyen fehérjéket multifunkcionálisnak nevezik.
3. A gyógyszerek ligandumok, amelyek befolyásolják a fehérjék működését. A fehérjék kölcsönhatása ligandumokkal specifikus. A fehérje és aktív centrumának konformációs labilitása miatt azonban lehetőség van egy másik olyan anyag kiválasztására, amely szintén kölcsönhatásba léphet a fehérjével az aktív centrumban vagy a molekula más részében.
A természetes ligandumhoz hasonló szerkezetű anyagot ún a ligandum szerkezeti analógja vagy természetellenes ligandum. Az aktív helyen lévő fehérjével is kölcsönhatásba lép. Egy ligandum szerkezeti analógja egyaránt fokozhatja a fehérje működését (agonista),és csökkentse azt (antagonista). A ligandum és szerkezeti analógjai versengenek egymással a fehérjéhez való kötődésért ugyanazon a helyen. Az ilyen anyagokat ún versenyképes modulátorok fehérjefunkciók (szabályozói). Sok gyógyszer fehérje inhibitorként működik. Ezek egy részét természetes ligandumok kémiai módosításával állítják elő. A fehérjefunkciók gátlói lehetnek gyógyszerek és mérgek.
Az atropin az M-kolinerg receptorok kompetitív inhibitora. Az acetilkolin egy neurotranszmitter az idegimpulzusok kolinerg szinapszisokon keresztül történő továbbítására. A gerjesztés végrehajtásához a szinaptikus hasadékba felszabaduló acetilkolinnak kölcsönhatásba kell lépnie a posztszinaptikus membrán receptorfehérjével. Két típust találtunk kolinerg receptorok:
M receptor az acetilkolinon kívül szelektív kölcsönhatásba lép a muszkarinnal (légyölő galóca toxin). M - kolinerg receptorok jelen vannak a simaizmokon, és az acetilkolinnal kölcsönhatásba lépve összehúzódásukat okozzák;
H receptor specifikusan kötődik a nikotinhoz. Az N-kolinerg receptorok a harántcsíkolt vázizmok szinapszisaiban találhatók.
Specifikus inhibitor M-kolinerg receptorok az atropin. Belladonna és tyúkszem növényekben található.
 Az atropin szerkezetében és térbeli elrendezésében az acetilkolinhoz hasonló funkcionális csoportokat tartalmaz, ezért az M-kolinerg receptorok kompetitív inhibitora. Tekintettel arra, hogy az acetilkolin M-kolinerg receptorokhoz való kötődése a simaizmok összehúzódását okozza, az atropint olyan gyógyszerként használják, amely enyhíti a görcsöket. (görcsoldó).Így ismert az atropin alkalmazása a szemizmok ellazítására a szemfenék megtekintésekor, valamint a gyomor-bélrendszeri kólika során fellépő görcsök enyhítésére. Az M-kolinerg receptorok a központi idegrendszerben (CNS) is jelen vannak, ezért nagy dózisú atropin nemkívánatos reakciót válthat ki a központi idegrendszerből: motoros és mentális izgatottságot, hallucinációkat, görcsöket.
Az atropin szerkezetében és térbeli elrendezésében az acetilkolinhoz hasonló funkcionális csoportokat tartalmaz, ezért az M-kolinerg receptorok kompetitív inhibitora. Tekintettel arra, hogy az acetilkolin M-kolinerg receptorokhoz való kötődése a simaizmok összehúzódását okozza, az atropint olyan gyógyszerként használják, amely enyhíti a görcsöket. (görcsoldó).Így ismert az atropin alkalmazása a szemizmok ellazítására a szemfenék megtekintésekor, valamint a gyomor-bélrendszeri kólika során fellépő görcsök enyhítésére. Az M-kolinerg receptorok a központi idegrendszerben (CNS) is jelen vannak, ezért nagy dózisú atropin nemkívánatos reakciót válthat ki a központi idegrendszerből: motoros és mentális izgatottságot, hallucinációkat, görcsöket.
A ditilin a H-kolinerg receptorok kompetitív agonistája, gátolja a neuromuszkuláris szinapszisok működését.
 A vázizmok neuromuszkuláris szinapszisai H-kolinerg receptorokat tartalmaznak. Az acetilkolinnal való kölcsönhatásuk izomösszehúzódásokhoz vezet. Egyes sebészeti beavatkozások során, valamint endoszkópos vizsgálatok során olyan gyógyszereket alkalmaznak, amelyek a vázizmok ellazulását okozzák. (izomrelaxánsok). Ide tartozik a ditilin, amely az acetilkolin szerkezeti analógja. H-kolinerg receptorokhoz kötődik, de az acetilkolintól eltérően az acetilkolinészteráz enzim nagyon lassan tönkreteszi. Az ioncsatornák elhúzódó megnyitása és a membrán tartós depolarizációja következtében az idegimpulzusok vezetése megszakad, és izomrelaxáció következik be. Kezdetben ezeket a tulajdonságokat a curare méregben fedezték fel, ezért nevezik az ilyen gyógyszereket curare-szerű.
A vázizmok neuromuszkuláris szinapszisai H-kolinerg receptorokat tartalmaznak. Az acetilkolinnal való kölcsönhatásuk izomösszehúzódásokhoz vezet. Egyes sebészeti beavatkozások során, valamint endoszkópos vizsgálatok során olyan gyógyszereket alkalmaznak, amelyek a vázizmok ellazulását okozzák. (izomrelaxánsok). Ide tartozik a ditilin, amely az acetilkolin szerkezeti analógja. H-kolinerg receptorokhoz kötődik, de az acetilkolintól eltérően az acetilkolinészteráz enzim nagyon lassan tönkreteszi. Az ioncsatornák elhúzódó megnyitása és a membrán tartós depolarizációja következtében az idegimpulzusok vezetése megszakad, és izomrelaxáció következik be. Kezdetben ezeket a tulajdonságokat a curare méregben fedezték fel, ezért nevezik az ilyen gyógyszereket curare-szerű.
TÉMAKÖR 1.3. A FEHÉRJÉK DENATURÁLÁSA ÉS SPONTÁN RENATIVÁLÁSUK LEHETŐSÉGE
1. Mivel a fehérjék natív konformációja a gyenge kölcsönhatások miatt megmarad, a fehérjét körülvevő környezet összetételének és tulajdonságainak változása, a kémiai reagenseknek és fizikai tényezőknek való kitettség változást okoz a konformációjukban (a konformációs labilitás tulajdonsága). Nagyszámú kötés felszakadása a fehérjék natív konformációjának megsemmisüléséhez és denaturálásához vezet.
A fehérjék denaturációja- ez a natív konformációjuk denaturáló szerek hatására bekövetkező pusztulása, amit a fehérje térszerkezetét stabilizáló gyenge kötések felszakadása okoz. A denaturáció a fehérje egyedi háromdimenziós szerkezetének és aktív centrumának tönkremenetelével és biológiai aktivitásának elvesztésével jár (1.16. ábra).
Egy fehérje minden denaturált molekulája véletlenszerű konformációt kap, amely különbözik ugyanazon fehérje többi molekulájától. Az aktív centrumot alkotó aminosavgyökök egymástól térben távolinak bizonyulnak, pl. a fehérje ligandummal való specifikus kötőhelye elpusztul. A denaturáció során a fehérjék elsődleges szerkezete változatlan marad.
Denaturáló szerek alkalmazása a biológiai kutatásban és az orvostudományban. A biokémiai vizsgálatok során a biológiai anyagokban lévő kis molekulatömegű vegyületek meghatározása előtt a fehérjéket általában először eltávolítják az oldatból. Erre a célra leggyakrabban triklór-ecetsavat (TCA) használnak. Miután TCA-t adtunk az oldathoz, a denaturált fehérjék kicsapódnak, és szűréssel könnyen eltávolíthatók (1.1. táblázat).
A gyógyászatban a denaturáló szereket gyakran használják orvosi műszerek és anyagok autoklávokban történő sterilizálására (a denaturálószer magas hőmérsékletű), valamint antiszeptikumként (alkohol, fenol, klóramin) a patogén mikroflórát tartalmazó szennyezett felületek kezelésére.
2. Spontán fehérje reaktiváció- a fehérjék elsődleges szerkezetének, konformációjának és működésének determinizmusának bizonyítása. Az egyes fehérjék egy gén termékei, amelyek azonos aminosav-szekvenciával rendelkeznek, és azonos konformációt kapnak a sejtben. Azt az alapvető következtetést, hogy a fehérje elsődleges szerkezete már tartalmaz információkat a konformációjáról és funkciójáról, néhány fehérje (különösen a ribonukleáz és a mioglobin) azon képessége alapján született, hogy spontán renativálódnak - denaturáció után visszaállítják natív konformációjukat.
A térbeli fehérjeszerkezetek kialakítása az önszerveződési módszerrel történik - egy spontán folyamat, amelyben egy egyedi primer szerkezetű polipeptidlánc hajlamos az oldatban a legalacsonyabb szabadenergiájú konformációt felvenni. A ribonukleáz enzimmel végzett kísérletben leírták a denaturáció után elsődleges szerkezetüket megőrző fehérjék renativálásának képességét.
A ribonukleáz egy enzim, amely lebontja az egyes nukleotidok közötti kötéseket egy RNS-molekulában. Ennek a globuláris fehérjének egy polipeptidlánca van, amelynek harmadlagos szerkezetét sok gyenge és négy diszulfidkötés stabilizálja.
A ribonukleáz karbamiddal, amely megszakítja a hidrogénkötéseket a molekulában, és egy redukálószerrel, amely megszakítja a diszulfid kötéseket, az enzim denaturálásához és aktivitásának elvesztéséhez vezet.
A denaturáló szerek dialízissel történő eltávolítása a fehérje konformációjának és funkciójának helyreállításához vezet, azaz. az újjászületésre. (1.17. ábra).
 Rizs. 1.17. A ribonukleáz denaturálása és renativációja
Rizs. 1.17. A ribonukleáz denaturálása és renativációja
A - a ribonukleáz natív konformációja, amelynek harmadlagos szerkezetében négy diszulfid kötés található; B - denaturált ribonukleáz molekula;
B - reaktivált ribonukleáz molekula helyreállított szerkezettel és funkcióval
1. Töltse ki az 1.2 táblázatot!
1.2. táblázat. Az aminosavak osztályozása a gyökök polaritása szerint
2. Írd fel a tetrapeptid képletét:
Asp - Pro - Fen - Liz
a) kiemeli a peptidben a peptidvázat alkotó ismétlődő csoportokat és az aminosavgyökök által képviselt változó csoportokat;
b) jelölje meg az N- és C-terminust;
c) kiemeljük a peptidkötéseket;
d) írjon egy másik peptidet, amely ugyanazokból az aminosavakból áll;
e) megszámolja a hasonló aminosav-összetételű tetrapeptid lehetséges variánsainak számát.
3. Magyarázza meg a fehérjék elsődleges szerkezetének szerepét az emlős neurohypophysis két szerkezetileg hasonló és evolúciósan közeli peptidhormon - az oxitocin és a vazopresszin - összehasonlító elemzésének példáján (1.3. táblázat).
1.3. táblázat. Az oxitocin és a vazopresszin felépítése és funkciói
 Ezért:
Ezért:
a) hasonlítsa össze két peptid összetételét és aminosavszekvenciáját;
b) találja meg a két peptid primer szerkezetének hasonlóságát és biológiai hatásuk hasonlóságát;
c) megtalálni a két peptid szerkezeti és funkciói közötti különbségeket;
d) következtetések levonása a peptidek primer szerkezetének a funkciójukra gyakorolt hatásáról.
4. Ismertesse a globuláris fehérjék konformációjának kialakulásának főbb szakaszait (másodlagos, harmadlagos szerkezetek, a szuperszekunder szerkezet fogalma). Jelölje meg a fehérjeszerkezetek kialakításában részt vevő kötések típusait! Mely aminosavgyökök vehetnek részt hidrofób kölcsönhatások, ionos, hidrogénkötések kialakításában.
Adj rá példákat.
5. Határozza meg a „fehérjék konformációs labilitása” fogalmát, jelölje meg létezésének okait és jelentőségét.
6. Bővítse ki a következő kifejezés jelentését: „A fehérjék működése a ligandummal való specifikus kölcsönhatáson alapul”, a kifejezések használatával és jelentésük magyarázatával: fehérje konformáció, aktív centrum, ligandum, komplementaritás, fehérje funkciója.
7. Egy példa segítségével magyarázza el, mik azok a domének, és mi a szerepük a fehérjék működésében.
ÖNELLENŐRZÉSI FELADATOK
1. Mérkőzés.
Funkcionális csoport az aminosavgyökben:
A. Karboxilcsoport B. Hidroxilcsoport C Guanidincsoport D. Tiolcsoport E. Aminocsoport
2. Válaszd ki a helyes válaszokat.
A poláris töltetlen gyököket tartalmazó aminosavak a következők:
A. Cis B. Asn
B. Glu G. Három
3. Válaszd ki a helyes válaszokat.
Aminosav gyökök:
A. Adja meg az elsődleges szerkezet specifitását B. Vegyen részt a harmadlagos struktúra kialakításában
B. A fehérje felszínén helyezkednek el, befolyásolják annak oldhatóságát D. Aktív centrumot alkotnak
D. Részt vesz a peptidkötések kialakításában
4. Válaszd ki a helyes válaszokat.
Az aminosav gyökök között hidrofób kölcsönhatások alakulhatnak ki:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Válaszd ki a helyes válaszokat.
Az aminosav gyökök között ionos kötések jöhetnek létre:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn ápr
6. Válaszd ki a helyes válaszokat.
Az aminosav gyökök között hidrogénkötések alakulhatnak ki:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Mérkőzés.
A fehérjeszerkezet kialakításában részt vevő kötés típusa:
A. Elsődleges szerkezet B. Másodlagos szerkezet
B. Harmadlagos szerkezet
D. Szuperszekunder szerkezet E. Konformáció.
1. Hidrogénkötések a peptidváz atomjai között
2. Gyenge kötések aminosav gyökök funkcionális csoportjai között
3. Az aminosavak α-amino- és α-karboxilcsoportjai közötti kötések
8. Válaszd ki a helyes válaszokat. Tripszin:
A. Proteolitikus enzim B. Két domént tartalmaz
B. Hidrolizálja a keményítőt
D. Az aktív hely a tartományok között található. D. Két polipeptidláncból áll.
9. Válaszd ki a helyes válaszokat. Atropin:
A. Neurotranszmitter
B. Az acetilkolin szerkezeti analógja
B. Kölcsönhatásba lép a H-kolinerg receptorokkal
D. Erősíti az idegimpulzusok vezetését a kolinerg szinapszisokon keresztül
D. M-kolinerg receptorok kompetitív inhibitora
10. Válassza ki a megfelelő állításokat. A fehérjékben:
V. Az elsődleges struktúra információkat tartalmaz az aktív helyének szerkezetéről
B. Az aktív központ az elsődleges szerkezet szintjén alakul ki
B. A konformációt kovalens kötések mereven rögzítik
D. Az aktív hely kölcsönhatásba léphet hasonló ligandumok csoportjával
a D fehérjék konformációs labilitása miatt. A környezet változásai befolyásolhatják az aktív fehérjék affinitását
központ a ligandum
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
ALAPVETŐ FELTÉTELEK ÉS FOGALMAK
1. Fehérje, polipeptid, aminosavak
2. Primer, szekunder, harmadlagos fehérjeszerkezetek
3. Konformáció, natív fehérje konformáció
4. Kovalens és gyenge kötések a fehérjében
5. Konformációs labilitás
6. Fehérje aktív hely
7. Ligandumok
8. Fehérje hajtogatás
9. A ligandumok szerkezeti analógjai
10. Domain fehérjék
11. Egyszerű és összetett fehérjék
12. Fehérjedenaturálás, denaturáló szerek
13. Fehérjék reaktiválása
Problémákat megoldani
„A fehérjék szerkezeti szerveződése és működésük alapja”
1. A fehérje - hemoglobin A (HbA) fő funkciója az oxigén szállítása a szövetekbe. Az emberi populációban ennek a fehérjének többféle, megváltozott tulajdonságaival és funkcióval rendelkező formája ismert – az úgynevezett abnormális hemoglobinok. Például a sarlósejtes analízisben (HbS) szenvedő betegek vörösvérsejtjeiben található hemoglobin S-ről azt találták, hogy alacsony az oxigén parciális nyomása mellett (mint a vénás vérben) alacsony oldhatóságú. Ez a fehérje aggregátumainak kialakulásához vezet. A fehérje funkcióját elveszíti, kicsapódik, a vörösvértestek pedig szabálytalan alakúvá válnak (egy részük sarló alakot formál), és a lépben a szokásosnál gyorsabban pusztulnak el. Ennek eredményeként sarlósejtes vérszegénység alakul ki.
A HbA primer szerkezetében az egyetlen különbséget a hemoglobin β-lánc N-terminális régiójában találtuk. Hasonlítsa össze a β-lánc N-terminális régióit, és mutassa meg, hogy a fehérje primer szerkezetében bekövetkezett változások hogyan befolyásolják tulajdonságait és funkcióit.
 Ezért:
Ezért:
a) írja le az aminosavak képleteit, amelyekben a HbA különbözik, és hasonlítsa össze ezen aminosavak tulajdonságait (polaritás, töltés).
b) következtetést levonni az oldhatóság csökkenésének és a szövetekbe történő oxigénszállítás zavarának okáról.
2. Az ábrán egy olyan fehérje szerkezetének diagramja látható, amely ligandumokkal (aktív centrummal) rendelkezik kötőközponttal. Magyarázza el, hogy a fehérje miért szelektív a ligandum kiválasztásában. Ezért:
a) ne feledje, mi a fehérje aktív központja, és vegye figyelembe a fehérje aktív centrumának szerkezetét az ábrán látható módon;
b) írja fel az aktív centrumot alkotó aminosavgyökök képleteit!
c) rajzoljunk egy ligandumot, amely specifikusan kölcsönhatásba léphet a fehérje aktív helyével. Jelölje meg rajta azokat a funkciós csoportokat, amelyek az aktív centrumot alkotó aminosavgyökökkel kötést tudnak kialakítani;
d) jelölje meg az aktív centrum ligandum- és aminosavgyökei között létrejövő kötések típusait;
e) magyarázza el, hogy a fehérje-ligandum kölcsönhatás sajátossága min alapul.
 3.
Az ábrán a fehérje és számos ligandum aktív helye látható.
3.
Az ábrán a fehérje és számos ligandum aktív helye látható.
Határozza meg, hogy melyik ligandum lép a legnagyobb valószínűséggel kölcsönhatásba a fehérje aktív helyével, és miért.
 Milyen típusú kötések keletkeznek a fehérje-ligandum komplex képződése során?
Milyen típusú kötések keletkeznek a fehérje-ligandum komplex képződése során?
4. A természetes fehérjeligandumok szerkezeti analógjai gyógyszerként használhatók a fehérjék aktivitásának módosítására.
Az acetilkolin a neuromuszkuláris szinapszisokban a gerjesztés átvitelének közvetítője. Amikor az acetilkolin kölcsönhatásba lép a fehérjékkel - a vázizmok posztszinaptikus membránjának receptoraival, az ioncsatornák megnyílnak, és izomösszehúzódás következik be. A ditilin egy olyan gyógyszer, amelyet bizonyos műtéteknél használnak az izmok ellazítására, mivel megzavarja az idegimpulzusok átvitelét a neuromuszkuláris szinapszisokon keresztül. Ismertesse a ditilin izomrelaxáns hatásmechanizmusát! Ezért:
a) írja fel az acetilkolin és a ditilin képleteit, és hasonlítsa össze szerkezetüket;
b) ismertesse a ditilin relaxáló hatásának mechanizmusát!
5. Egyes betegségekben a páciens testhőmérséklete megemelkedik, amit a szervezet védekező reakciójának tekintenek. A magas hőmérséklet azonban káros a testfehérjékre. Magyarázza meg, hogy 40 °C feletti hőmérsékleten miért zavar a fehérjék működése, és miért fenyegeti az emberi életet! Ehhez ne feledje:
1) A fehérjék szerkezete és a szerkezetét a natív konformációban tartó kötések;
2) Hogyan változik a fehérjék szerkezete és működése a hőmérséklet emelkedésével?;
3) Mi a homeosztázis és miért fontos az emberi egészség megőrzése szempontjából?
2. moduláris egység OLIGOMERICK FEHÉRJEK MINT A SZABÁLYOZÁSI BEFOLYÁSOK CÉLJA. A FEHÉRJÉK SZERKEZETI ÉS FUNKCIÓS VÁLTOZÁSA. A FEHÉRJÉK ELVÁLASZTÁSÁNAK ÉS TISZTÍTÁSÁNAK MÓDSZEREI
Tanulási célok Legyen képes:
1. Az oligomer fehérjék szerkezetének és funkcióinak sajátosságaira vonatkozó ismeretek felhasználása funkcióik szabályozásának adaptív mechanizmusainak megértéséhez.
2. Ismertesse a chaperonok szerepét a fehérje konformáció szintézisében és fenntartásában celluláris körülmények között.
3. Magyarázza meg az élet megnyilvánulásainak sokféleségét a szervezetben szintetizálódó fehérjék szerkezetének és funkcióinak sokféleségével!
4. Elemezze a fehérjék szerkezete és funkciójuk közötti összefüggést rokon hemoproteinek - mioglobin és hemoglobin, valamint az immunglobulincsalád öt fehérjeosztályának képviselői - összehasonlításának példái segítségével.
5. Alkalmazza a fehérjék fiziko-kémiai tulajdonságainak sajátosságaira vonatkozó ismereteket más fehérjéktől és szennyeződésektől való tisztításukra.
6. A vérplazmafehérjék mennyiségi és minőségi összetételének eredményeinek értelmezése a klinikai diagnózis megerősítése vagy tisztázása érdekében.
Tud:
1. Az oligomer fehérjék szerkezetének jellemzői és funkciójuk szabályozásának adaptív mechanizmusai a hemoglobin példáján.
2. A chaperonok szerkezete és funkciói, valamint szerepük a fehérjék natív konformációjának fenntartásában celluláris körülmények között.
3. A fehérjék konformációjuk és funkcióik hasonlóságán alapuló családokba vonásának elvei immunglobulinok példáján.
4. A fehérjék fizikokémiai tulajdonságaik jellemzői alapján történő elválasztásának módszerei.
5. A vérplazma elektroforézise, mint a fehérjék minőségi és mennyiségi összetételének felmérésére szolgáló módszer.
TÉMAKÖR 1.4. AZ OLIGOMER FEHÉRJEK SZERKEZETÉNEK ÉS MŰKÖDÉSÉNEK JELLEMZŐI HEMOGLOBIN PÉLDÁJÁN
1. Sok fehérje több polipeptid láncot tartalmaz. Az ilyen fehérjéket ún oligomer,és az egyes láncok - protomerek. Az oligomer fehérjékben lévő protomereket sok gyenge, nem kovalens kötés köti össze (hidrofób, ionos, hidrogén). Kölcsönhatás
protomereket hajtanak végre, köszönhetően komplementaritásérintkező felületeik.
Az oligomer fehérjékben a protomerek száma nagyon eltérő lehet: a hemoglobin 4, az aszpartát-aminotranszferáz enzim 12, a dohánymozaikvírus fehérje pedig 2120 nem kovalens kötéssel összekapcsolt protomert tartalmaz. Következésképpen az oligomer fehérjék nagyon nagy molekulatömegűek lehetnek.
Egy protomer kölcsönhatása másokkal a fehérje-ligandum kölcsönhatás speciális esetének tekinthető, mivel mindegyik protomer ligandumként szolgál más protomerek számára. A protomerek számát és módját egy fehérjében ún kvaterner fehérjeszerkezet.
A fehérjék tartalmazhatnak azonos vagy eltérő szerkezetű protomereket, például a homodimerek két azonos protomert tartalmazó fehérjék, a heterodimerek pedig két különböző protomert tartalmazó fehérjék.
Ha a fehérjék különböző protomereket tartalmaznak, akkor rajtuk különböző ligandumokkal, szerkezetükben eltérő kötőcentrumok képződhetnek. Amikor egy ligandum kötődik az aktív helyhez, ennek a fehérjének a funkciója megnyilvánul. Egy másik protomeren elhelyezkedő központot allosztérikusnak nevezzük (az aktívtól eltérő). Kapcsolatfelvétel alloszterikus ligandum vagy effektor, szabályozó funkciót lát el (1.18. ábra). Az alloszterikus centrum kölcsönhatása az effektorral a teljes oligomer fehérje szerkezetében a konformációs labilitás miatt konformációs változásokat okoz. Ez befolyásolja az aktív hely affinitását egy specifikus ligandumhoz, és szabályozza a fehérje működését. Az összes protomer konformációjában és működésében bekövetkező változást egy oligomer fehérje és legalább egy ligandum kölcsönhatása során kooperatív konformációs változásoknak nevezzük. A fehérje működését fokozó effektorokat ún aktivátorok,és a működését gátló effektorok azok inhibitorok.
Így az oligomer fehérjék, valamint a doménszerkezettel rendelkező fehérjék a monomer fehérjékhez képest új tulajdonsággal rendelkeznek - a funkciók alloszterikus szabályozásának képessége (szabályozás különböző ligandumok fehérjéhez kapcsolásával). Ez látható két szorosan összefüggő komplex fehérje, a mioglobin és a hemoglobin szerkezetének és funkcióinak összehasonlításával.
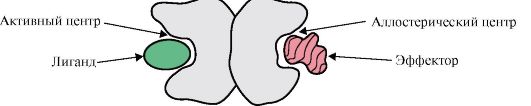 Rizs. 1.18. Egy dimer fehérje szerkezetének vázlata
Rizs. 1.18. Egy dimer fehérje szerkezetének vázlata
2. A mioglobin térszerkezetének kialakulása és működése.
A mioglobin (Mb) a vörös izmokban található fehérje, melynek fő feladata az intenzív izommunkához szükséges O 2 tartalékok létrehozása. Az Mb egy komplex fehérje, amely egy fehérjerészt - apoMb-t és egy nem fehérjerészt - hemet tartalmaz. Az apoMB elsődleges szerkezete határozza meg kompakt gömb alakú konformációját és az aktív centrum szerkezetét, amelyhez a mioglobin nem fehérje része, a hem kapcsolódik. A vérből az izmokba érkező oxigén a mioglobin Fe+ 2 heméhez kötődik. Az Mb egy monomer fehérje, amelynek nagyon nagy affinitása az O 2 iránt, ezért a mioglobin általi oxigén felszabadulás csak intenzív izommunka során történik, amikor az O 2 parciális nyomása meredeken csökken.
Az Mv konformáció kialakulása. A vörös izmokban a riboszómákon a transzláció során az MB elsődleges szerkezete szintetizálódik, amelyet egy 153 aminosavból álló specifikus szekvencia képvisel. Az Mb másodlagos szerkezete nyolc α-hélixet tartalmaz, amelyeket latin betűkkel A-tól H-ig neveznek, és amelyek között nem helikális régiók találhatók. Az Mb harmadlagos szerkezete kompakt gömb alakú, melynek mélyedésében az aktív centrum az F és az E α-hélix között helyezkedik el (1.19. ábra).
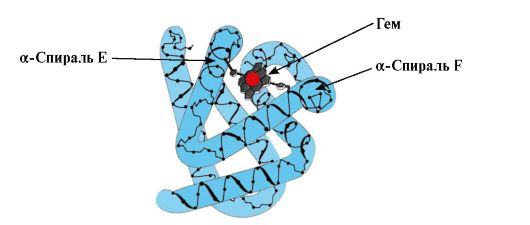 Rizs. 1.19. A mioglobin szerkezete
Rizs. 1.19. A mioglobin szerkezete
3. Az MV aktív központjának felépítésének és működésének jellemzői. Az Mb aktív centrumát túlnyomórészt hidrofób aminosav gyökök alkotják, amelyek az elsődleges szerkezetben egymástól távol helyezkednek el (például Tri 3 9 és Fen 138) Vízben rosszul oldódó ligandumok - hem és O 2 - kapcsolódnak az aktív centrumhoz. A hem az apoMB specifikus liganduma (1.20. ábra), melynek alapját négy pirrolgyűrű alkotja, amelyeket metenilhidak kötnek össze; a központban egy Fe+ 2 atom található, amely négy koordinációs kötéssel kapcsolódik a pirrolgyűrűk nitrogénatomjaihoz. Az Mb aktív centrumában a hidrofób aminosavgyökök mellett két aminosav aminosavmaradéka is található hidrofil gyökökkel - Gis E 7(Gis 64) és GIS F 8(Ő 93) (1.21. ábra).
 Rizs. 1.20. A hem szerkezete a mioglobin és a hemoglobin nem fehérje része
Rizs. 1.20. A hem szerkezete a mioglobin és a hemoglobin nem fehérje része
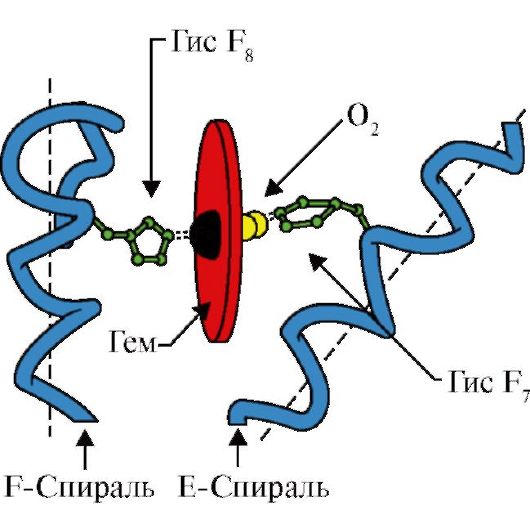 Rizs. 1.21. A hem és az O2 elhelyezkedése az apomioglobin és a hemoglobin protomerek aktív helyén
Rizs. 1.21. A hem és az O2 elhelyezkedése az apomioglobin és a hemoglobin protomerek aktív helyén
A hem kovalensen kötődik az F8-hoz egy vasatomon keresztül. Az O 2 a hemsík másik oldalán tapad a vashoz. Az ő E 7 szükséges az O 2 helyes orientációjához, és elősegíti az oxigén hozzáadását a Fe + 2 hemhez
GIS F 8 koordinációs kötést képez a Fe+ 2-vel, és szilárdan rögzíti a hemet az aktív központban. Gis E 7 szükséges a helyes orientációhoz egy másik ligandum aktív központjában - O 2 a Fe + 2 hem kölcsönhatása során. A hem mikrokörnyezete megteremti a feltételeket az O 2 erős, de reverzibilis Fe + 2-hez való kötéséhez, és megakadályozza a víz bejutását a hidrofób aktív helyre, ami Fe + 3 -dá oxidációjához vezethet.
Az Mb monomer szerkezete és aktív centruma meghatározza a fehérje O 2 iránti nagy affinitását.
4. A Hb oligomer szerkezete és a Hb O 2 ligandumokhoz való affinitásának szabályozása. Humán hemoglobinok- olyan fehérjék családja, mint a mioglobin, amely komplex fehérjékhez (hemoproteinekhez) kapcsolódik. Tetramer szerkezetűek és két α-láncot tartalmaznak, de a másik két polipeptid lánc (2α-, 2x-láncok) szerkezetében különböznek. A második polipeptidlánc szerkezete meghatározza a Hb ezen formáinak működésének sajátosságait. A felnőttek vörösvérsejtjeiben a hemoglobin körülbelül 98%-a hemoglobin A(2α-, 2p-láncok).
A magzati fejlődés során a hemoglobin két fő típusa működik: embrionális Hb(2α, 2ε), amely a magzati fejlődés korai szakaszában található, ill hemoglobin F (magzati)- (2α, 2γ), amely a méhen belüli fejlődés hatodik hónapjában helyettesíti a korai magzati hemoglobint, és csak a születés után váltja fel Hb A-val.
A HB A a felnőtt emberi vörösvértestekben található mioglobinhoz (MB) rokon fehérje. Egyedi protomereinek szerkezete hasonló a mioglobinéhoz. A mioglobin és a hemoglobin protomerek másodlagos és harmadlagos szerkezete nagyon hasonló, annak ellenére, hogy polipeptid láncaik elsődleges szerkezetében mindössze 24 aminosav azonos (a hemoglobin protomereinek másodlagos szerkezete a mioglobinhoz hasonlóan nyolc α-hélixet tartalmaz, latin betűkkel jelölve A-tól H -ig, és a harmadlagos szerkezet tömör gömb alakú). A mioglobintól eltérően azonban a hemoglobin oligomer szerkezetű, négy polipeptidláncból áll, amelyeket nem kovalens kötés köt össze (1.22. ábra).
Minden Hb protomer egy nem fehérje részhez kapcsolódik - a hemhez és a szomszédos protomerekhez. A Hb fehérje részének kapcsolódása a hemhez hasonló a mioglobinhoz: a fehérje aktív centrumában a hem hidrofób részeit hidrofób aminosavgyökök veszik körül, a His F 8 és His E 7 kivételével, amelyek a hem síkjának két oldalán helyezkednek el és hasonló szerepet játszanak a fehérje működésében és oxigénnel való megkötésében (lásd a mioglobin szerkezetét).
 Rizs. 1.22. A hemoglobin oligomer szerkezete
Rizs. 1.22. A hemoglobin oligomer szerkezete
Kívül, Gis E 7 fontosat végez további szerep az Nv működésében. A szabad hem 25 000-szer nagyobb affinitást mutat a CO-hoz, mint az O2-hoz. A CO kis mennyiségben képződik a szervezetben, és a hem iránti nagy affinitása miatt megzavarhatja a sejtek életéhez szükséges O 2 szállítását. A hemoglobin összetételében azonban a hem szén-monoxidhoz való affinitása csak 200-szor haladja meg az O 2 affinitását, mivel a His E 7 az aktív centrumban van jelen. Ennek az aminosavnak a fennmaradó része optimális feltételeket teremt a hem O 2-hez való kötődéséhez, és gyengíti a hem és a CO kölcsönhatását.
5. A HB fő funkciója az O2 szállítása a tüdőből a szövetekbe. Ellentétben a monomer mioglobinnal, amelynek nagyon nagy affinitása van az O2-hoz, és az oxigén tárolását végzi a vörös izmokban, a hemoglobin oligomer szerkezete biztosítja:
1) a HB gyors telítődése oxigénnel a tüdőben;
2) a HB azon képessége, hogy oxigént szabadít fel a szövetekben viszonylag magas O 2 parciális nyomáson (20-40 Hgmm);
3) a Hb O 2-hez való affinitásának szabályozásának lehetősége.
6. A hemoglobin protomerek konformációjának kooperatív változásai felgyorsítják az O 2 megkötését a tüdőben és a szövetekbe való kibocsátását. A tüdőben az O 2 magas parciális nyomása elősegíti a Hb kötődését négy protomer (2α és 2β) aktív helyén. Minden protomer aktív központja, akárcsak a mioglobinban, két α-hélix (F és E) között helyezkedik el egy hidrofób zsebben. Tartalmaz egy nem fehérje részt - hem, amely sok gyenge hidrofób kölcsönhatás révén kapcsolódik a fehérje részhez, valamint egy erős kötés a Fe 2 + hem és a His F 8 között (lásd 1.21. ábra).
A dezoxihemoglobinban ennek a His F 8-hoz való kötődésnek köszönhetően a Fe 2 + atom a hem síkjából a hisztidin felé nyúlik ki. Az O 2 kötődése Fe 2 + -hoz a hem másik oldalán, a His E 7 régióban, egyetlen szabad koordinációs kötés segítségével történik. His E 7 optimális feltételeket biztosít az O 2 hemvashoz való kötődéséhez.
O 2 hozzáadása az egyik protomer Fe + 2 atomjához a hem síkba való mozgását okozza, amit a hozzá kapcsolódó hisztidincsoport követ.
 Rizs. 1.23. A hemoglobin protomer konformációjának változása O 2 -vel kombinálva
Rizs. 1.23. A hemoglobin protomer konformációjának változása O 2 -vel kombinálva
Ez az összes polipeptidlánc konformációjának megváltozásához vezet a konformációs labilitásuk miatt. Más láncok konformációjának megváltoztatása megkönnyíti kölcsönhatásukat a következő O 2 molekulákkal.
A negyedik O 2 molekula 300-szor könnyebben kötődik a hemoglobinhoz, mint az első (1.24. ábra).
 Rizs. 1.24. A hemoglobin protomerek konformációjának kooperatív változásai az O2-val való kölcsönhatás során
Rizs. 1.24. A hemoglobin protomerek konformációjának kooperatív változásai az O2-val való kölcsönhatás során
A szövetekben minden következő O 2 molekula könnyebben hasad le, mint az előző, a protomerek konformációjában bekövetkező kooperatív változások miatt is.
7. A szerves anyagok lebontása során képződő CO 2 és H+ koncentrációjuk arányában csökkenti a hemoglobin O 2 iránti affinitását. A sejtek működéséhez szükséges energia elsősorban a mitokondriumokban termelődik a szerves anyagok oxidációja során, a tüdőből a hemoglobin által szállított O 2 felhasználásával. A szerves anyagok oxidációja következtében bomlásuk végtermékei keletkeznek: CO 2 és K 2 O, amelyek mennyisége arányos a folyamatban lévő oxidációs folyamatok intenzitásával.
A CO 2 a sejtekből a vérbe diffundál, és behatol a vörösvérsejtekbe, ahol a karbanhidráz enzim hatására szénsavvá alakul. Ez a gyenge sav protonra és bikarbonát ionra disszociál.
A H+ képes csatlakozni az Ő radikálisaihoz 14 6 a hemoglobin α- és β-láncában, azaz. a hemtől távoli területeken. A hemoglobin protonálása csökkenti az O 2 iránti affinitását, elősegíti az O 2 eltávolítását az oxiHb-ből, a dezoxiHb képződését, és a képződött protonok számával arányosan növeli a szövetek oxigénellátását (1.25. ábra).
A felszabaduló oxigén mennyiségének a vörösvérsejtek H+-koncentrációjának növekedésétől függő növekedését Bohr-effektusnak nevezik (a dán fiziológus, Christian Bohr után kapta a nevét, aki először fedezte fel ezt a hatást).
A tüdőben az oxigén nagy parciális nyomása elősegíti annak dezoxiHb-hez való kötődését, ami csökkenti a fehérje H + iránti affinitását. A szénsav hatására felszabaduló protonok reakcióba lépnek hidrogén-karbonátokkal, így CO 2 és H 2 O keletkezik.

 Rizs. 1.25. A Hb O 2 iránti affinitásának függése a CO 2 és a protonok koncentrációjától (Bohr-effektus):
Rizs. 1.25. A Hb O 2 iránti affinitásának függése a CO 2 és a protonok koncentrációjától (Bohr-effektus):
A- a CO 2 és H+ koncentráció hatása az O 2 felszabadulására a HB komplexből (Bohr-effektus); B- a dezoxihemoglobin oxigenizációja a tüdőben, CO 2 képződése és felszabadulása.
A keletkező CO 2 az alveoláris térbe kerül, és a kilélegzett levegővel távozik. Így a hemoglobin által a szövetekben felszabaduló oxigén mennyiségét a szerves anyagok lebontásának termékei szabályozzák: minél intenzívebb az anyagok lebomlása, például a testmozgás során, annál nagyobb a CO 2 és H + koncentrációja, és annál több az oxigén. a szövetek a Hb O 2 iránti affinitásának csökkenése következtében kapnak.
8. A Hb O2 iránti affinitásának alloszterikus szabályozása a ligandum - 2,3-biszfoszfoglicerát - által. Az eritrocitákban a hemoglobin allosztérikus liganduma, a 2,3-biszfoszfoglicerát (2,3-BPG) szintetizálódik a glükóz oxidációjának termékéből - 1,3-biszfoszfoglicerátból. Normál körülmények között a 2,3-BPG koncentrációja magas, és összehasonlítható a Hb koncentrációjával. A 2,3-BPG erős negatív töltése -5.
 A szöveti kapillárisokban lévő biszfoszfoglicerát, amely a dezoxihemoglobinhoz kötődik, növeli az oxigén felszabadulását a szövetekbe, csökkentve a Hb O 2 iránti affinitását.
A szöveti kapillárisokban lévő biszfoszfoglicerát, amely a dezoxihemoglobinhoz kötődik, növeli az oxigén felszabadulását a szövetekbe, csökkentve a Hb O 2 iránti affinitását.
A tetramer hemoglobinmolekula közepén egy üreg található. Mind a négy protomer aminosav-maradékai alkotják (lásd 1.22. ábra). A szöveti kapillárisokban a Hb protonálódása (Bohr-effektus) a hem vas és az O2 közötti kötés megszakadásához vezet. Egy molekulában
dezoxihemoglobin, az oxihemoglobinhoz képest további ionos kötések jelennek meg, amelyek összekötik a protomereket, aminek következtében a központi üreg méretei megnőnek az oxihemoglobinhoz képest. A központi üreg az a hely, ahol a 2,3-BPG a hemoglobinhoz kötődik. A központi üreg méretének különbsége miatt a 2,3-BPG csak a dezoxihemoglobinhoz tud kapcsolódni.
A 2,3-BPG kölcsönhatásba lép a hemoglobinnal a fehérje aktív központjaitól távoli helyen, és a allosztérikus(szabályozó) ligandumok, a Hb központi ürege pedig az allosztérikus központ. A 2,3-BPG erős negatív töltésű, és kölcsönhatásba lép a Hb két β-láncának öt pozitív töltésű csoportjával: a Val N-terminális α-aminocsoportjával és a Lys 82 His 143 gyökeivel (1.26. ábra).
 Rizs. 1.26. BPG a dezoxihemoglobin központi üregében
Rizs. 1.26. BPG a dezoxihemoglobin központi üregében
A BPG minden β-szálon három pozitív töltésű csoporthoz kötődik.
A szöveti kapillárisokban a keletkező dezoxihemoglobin kölcsönhatásba lép a 2,3-BPG-vel, és ionos kötések jönnek létre a β-láncok pozitív töltésű gyökök és a negatív töltésű ligandum között, amelyek megváltoztatják a fehérje konformációját és csökkentik a Hb affinitását az oxigénhez. . A Hb O 2 iránti affinitásának csökkenése hozzájárul az O 2 hatékonyabb felszabadulásához a szövetekbe.
A tüdőben nagy parciális nyomáson az oxigén kölcsönhatásba lép a Hb-vel, csatlakozik a hem vashoz; Ebben az esetben a fehérje konformációja megváltozik, a központi üreg csökken, és a 2,3-BPG kiszorul az allosztérikus centrumból
Így az oligomer fehérjék új tulajdonságokkal rendelkeznek a monomer fehérjékhez képest. Ligandumok kötődése a helyeken
egymástól térben távol eső (allosztérikus), konformációs változásokat okozhat a teljes fehérjemolekulában. A szabályozó ligandumokkal való kölcsönhatás következtében a fehérjemolekula konformációja megváltozik, és funkciója alkalmazkodik a környezeti változásokhoz.
TÉMAKÖR 1.5. A FEHÉRJÉK NATÍV KONFORMÁCIÓJÁNAK FENNTARTÁSA SEJTKÖRÜLMÉNYEK KÖZÖTT
A sejtekben a polipeptid láncok szintézise során a membránokon keresztül a sejt megfelelő részeibe történő szállítása során a folding folyamata (a natív konformáció kialakulása) és az oligomer fehérjék összerakása során, valamint működésük során, intermedier , aggregációra hajlamos, instabil konformációk keletkeznek a fehérje szerkezetében. A hidrofób gyökök, amelyek általában a natív konformációban a fehérjemolekulában rejtőznek, instabil konformációban jelennek meg a felszínen, és hajlamosak más, vízben rosszul oldódó fehérjék csoportjaival kombinálódni. Valamennyi ismert élőlény sejtjében olyan speciális fehérjéket találtak, amelyek biztosítják a sejtfehérjék optimális feltekeredését, működés közben stabilizálják natív konformációjukat, és ami a legfontosabb, a homeosztázis zavara esetén fenntartják az intracelluláris fehérjék szerkezetét és funkcióit. Ezeket a fehérjéket ún "kísérők" ami franciául „dadát” jelent.
1. Molekuláris chaperonok és szerepük a fehérjedenaturáció megelőzésében.
A chaperonokat (CH) alegységeik tömege szerint osztályozzák. A nagy molekulatömegű chaperonok tömege 60-110 kDa. Közülük három osztályt vizsgáltak a legtöbbet: Sh-60, Sh-70 és Sh-90. Mindegyik osztály a rokon fehérjék családját tartalmazza. Így az Sh-70 66-78 kDa molekulatömegű fehérjéket tartalmaz. Az alacsony molekulatömegű chaperonok molekulatömege 40-15 kDa.
A kísérők között vannak alkotó fehérjék, amelyek magas szintű alapszintézise nem függ a szervezet sejtjeit érő stresszhatásoktól, és indukálható, amelynek szintézise normál körülmények között gyenge, de stressz hatására meredeken megnövekszik. Az indukálható chaperonokat „hősokk-fehérjéknek” is nevezik, mivel először magas hőmérsékletnek kitett sejtekben fedezték fel őket. A sejtekben a fehérjék magas koncentrációja miatt a részben denaturált fehérjék spontán reaktiválása nehézkes. Az Sh-70 megakadályozhatja a denaturáció kialakulását és segíthet a fehérjék natív konformációjának helyreállításában. Molekuláris chaperonok-70- a fehérjék egy erősen konzervált osztálya, amely a sejt minden részében megtalálható: citoplazmában, sejtmagban, endoplazmatikus retikulumban, mitokondriumokban. Az egyetlen polipeptidlánc Ш-70 karboxil végén van egy olyan régió, amely egy barázda, amely képes kölcsönhatásba lépni hosszúságú peptidekkel.
7-9 aminosavból, hidrofób gyökökkel dúsítva. Az ilyen régiók a globuláris fehérjékben körülbelül minden 16 aminosavban előfordulnak. Az Sh-70 képes megvédeni a fehérjéket a hőmérsékleti inaktivációtól, és helyreállítani a részlegesen denaturált fehérjék konformációját és aktivitását.
2. A chaperonok szerepe a fehérje feltekeredésében. A riboszómán történő fehérjeszintézis során a polipeptid N-terminális régiója szintetizálódik a C-terminális előtt. A natív konformáció kialakításához a fehérje teljes aminosavszekvenciája szükséges. A fehérjeszintézis folyamatában a chaperon-70 az aktív centrum szerkezetéből adódóan a szintézis befejezéséig képesek lezárni a polipeptid aggregációra hajlamos, hidrofób aminosavgyökökben dúsított területeit (1.27. ábra, A) ).
 Rizs. 1.27. A chaperonok részvétele a fehérje hajtogatásban
Rizs. 1.27. A chaperonok részvétele a fehérje hajtogatásban
A - chaperon-70 részvétele a szintetizált polipeptid szakaszai közötti hidrofób kölcsönhatások megelőzésében; B - a fehérje natív konformációjának kialakulása a chaperon komplexben
Sok nagy molekulájú fehérje, amelyek összetett konformációval, például doménszerkezettel rendelkeznek, az Sh-60 által alkotott speciális térben hajtódnak össze. Ш-60 14 alegységből álló oligomer komplexként működik. Két üreges gyűrűt alkotnak, amelyek mindegyike hét alegységből áll, ezek a gyűrűk kapcsolódnak egymáshoz. Mindegyik Sh-60 alegység három doménből áll: apikális (apikális), hidrofób gyökökkel dúsítva, amelyek a gyűrű ürege felé néznek, intermedier és ekvatoriális (1.28. ábra).
 Rizs. 1.28. A 14 Ш-60-ból álló chaperonin komplex felépítése
Rizs. 1.28. A 14 Ш-60-ból álló chaperonin komplex felépítése
A - oldalnézet; B - felülnézet
A szintetizált fehérjék, amelyek felületén a kibontott molekulákra jellemző elemek, különösen hidrofób gyökök vannak, bejutnak a chaperongyűrűk üregébe. Ezeknek az üregeknek a sajátos környezetében addig kutatjuk a lehetséges konformációkat, amíg meg nem találjuk az egyetlen, amely energetikailag a legkedvezőbb (1.27. ábra, B). A konformációk kialakulását és a fehérje felszabadulását az ekvatoriális régióban ATP hidrolízis kíséri. Jellemzően az ilyen chaperone-függő hajtogatás jelentős mennyiségű energiát igényel.
Amellett, hogy részt vesznek a fehérjék háromdimenziós szerkezetének kialakításában és a részben denaturált fehérjék renativációjában, a chaperonok olyan alapvető folyamatok lezajlásához is szükségesek, mint az oligomer fehérjék összeállítása, a denaturált fehérjék felismerése és lizoszómákba történő szállítása, fehérjék membránokon keresztül történő transzportja, és részvétel a fehérjekomplexek aktivitásának szabályozásában.
TÉMAKÖR 1.6. VÁLTOZATOS FEHÉRJÉK. FEHÉRJECSALÁDOK IMUNOGLOBULINOK PÉLDÁJÁN
1. A fehérjék az egyes sejtek és az egész többsejtű szervezet életében meghatározó szerepet töltenek be, funkciójuk pedig meglepően sokrétű. Ezt a fehérjék elsődleges szerkezetének és konformációinak jellemzői, az aktív centrum egyedi szerkezete és a specifikus ligandumok megkötő képessége határozzák meg.
A peptidláncok összes lehetséges változatának csak nagyon kis része tud stabil térbeli szerkezetet felvenni; többség
ezek közül sok konformációt vehet fel megközelítőleg azonos Gibbs-energiával, de eltérő tulajdonságokkal. A legtöbb ismert fehérje biológiai evolúcióval kiválasztott elsődleges szerkezete biztosítja az egyik konformáció kivételes stabilitását, amely meghatározza e fehérje működésének jellemzőit.
2. Fehérjecsaládok. Ugyanazon biológiai fajon belül az aminosavak szubsztitúciói különböző fehérjék megjelenéséhez vezethetnek, amelyek rokon funkciókat látnak el, és homológ aminosavszekvenciákkal rendelkeznek. Az ilyen rokon fehérjék feltűnően hasonló konformációkkal rendelkeznek: az α-hélixek és/vagy β-struktúrák száma és relatív helyzete, a polipeptidláncok legtöbb fordulata és hajlata hasonló vagy azonos. A polipeptidlánc homológ régióit, hasonló konformációt és kapcsolódó funkciókat tartalmazó fehérjéket fehérjecsaládokba sorolják. Példák fehérjecsaládokra: szerin proteinázok, immunglobulin család, mioglobin család.
Szerin proteinázok- fehérjecsalád, amely proteolitikus enzimek funkcióját látja el. Ide tartoznak az emésztőenzimek - kimotripszin, tripszin, elasztáz és számos véralvadási faktor. Ezek a fehérjék pozíciójuk 40%-ában azonos aminosavakat és nagyon hasonló konformációt tartalmaznak (1.29. ábra).

Rizs. 1.29. Az elasztáz (A) és a kimotripszin (B) térbeli szerkezete
Egyes aminosav-szubsztitúciók változásokhoz vezettek e fehérjék szubsztrát-specifitásában, és funkcionális diverzitás kialakulásához vezettek a családon belül.
3. Immunglobulin család. Az immunrendszer működésében óriási szerepet játszanak az immunglobulin szupercsalád fehérjéi, amely három fehérjecsaládot foglal magában:
antitestek (immunglobulinok);
T-limfocita receptorok;
A fő hisztokompatibilitási komplex fehérjéi – MHC 1. és 2. osztály (Major Histocompatibility Complex).
Mindezek a fehérjék doménszerkezettel rendelkeznek, homológ immunszerű doménekből állnak, és hasonló funkciókat látnak el: kölcsönhatásba lépnek idegen struktúrákkal, akár a vérben, nyirok- vagy sejtközi folyadékban (antitestek) oldva, akár a sejtek felszínén (saját ill. külföldi).
4. Antitestek- a B-limfociták által termelt specifikus fehérjék egy idegen struktúra szervezetbe jutására válaszul, ún antigén.
Az antitestek szerkezetének jellemzői
A legegyszerűbb antitestmolekulák négy polipeptid láncból állnak: két egyforma könnyű - L, amely körülbelül 220 aminosavat tartalmaz, és két azonos nehéz - H, amely 440-700 aminosavból áll. Az antitestmolekulában mind a négy láncot sok nem kovalens kötés és négy diszulfidkötés köti össze (1.30. ábra).
Az antitest könnyű láncai két doménből állnak: egy variábilis doménből (VL), amely a polipeptidlánc N-terminális régiójában található, és egy konstans doménből (CL), amely a C-terminálison található. A nehézláncoknak általában négy doménje van: egy változó (VH), amely az N-terminálison található, és három konstans domén (CH1, CH2, CH3) (lásd 1.30. ábra). Mindegyik immunglobulin doménnek van egy β-lemez szuperstruktúrája, amelyben két ciszteinmaradék diszulfidkötéssel kapcsolódik.
A két konstans CH1 és CH2 domén között van egy nagyszámú prolinmaradékot tartalmazó régió, amely megakadályozza a másodlagos szerkezet kialakulását és a szomszédos H-láncok kölcsönhatását ebben a szegmensben. Ez a csuklórégió rugalmasságot biztosít az antitestmolekulának. A nehéz és könnyű láncok variábilis doménjei között két azonos antigénkötő hely (aktív antigénkötő hely) található, ezért az ilyen antitesteket gyakran ún. bivalensek. Az antigén antitesthez való kötődésében nem mindkét lánc variábilis régióinak teljes aminosavszekvenciája vesz részt, hanem csak 20-30 aminosav, amely az egyes láncok hipervariábilis régióiban található. Ezek a régiók határozzák meg az egyes típusú antitestek egyedi képességét a megfelelő komplementer antigénnel való kölcsönhatásra.
Az antitestek jelentik a szervezet egyik védelmi vonalát a behatoló idegen organizmusok ellen. Működésük két szakaszra osztható: az első szakasz az antigén felismerése és megkötése idegen organizmusok felszínén, ami az antitest-szerkezetben található antigénkötő helyek miatt lehetséges; a második szakasz az antigén inaktiválási és elpusztítási folyamatának beindítása. A második szakasz specifitása az antitestek osztályától függ. A nehéz láncoknak öt osztálya különbözik egymástól a konstans domének szerkezetében: α, δ, ε, γ és μ, amelyek szerint az immunglobulinok öt osztályát különböztetik meg: A, D, E, G és M.
A nehéz láncok szerkezeti jellemzői az egyes osztályokra jellemző konformációt adnak a nehéz láncok csuklórégióinak és C-terminális régióinak. Miután az antigén egy antitesthez kötődik, az állandó domének konformációs változásai határozzák meg az antigén eltávolításának útvonalát.
 Rizs. 1. 30. Az IgG doménszerkezete
Rizs. 1. 30. Az IgG doménszerkezete
Immunglobulinok M
Az M immunglobulinoknak két formája van.
Monomer forma- A fejlődő B-limfociták által termelt antitestek 1. osztálya. Ezt követően sok B-sejt átáll más osztályú antitestek termelésére, de ugyanazzal az antigénkötő hellyel. Az IgM a membránba van ágyazva, és antigénfelismerő receptorként működik. Az IgM beépülése a sejtmembránba a farokrégióban 25 hidrofób aminosav jelenléte miatt lehetséges.
Az IgM szekréciós formájaöt monomer alegységet tartalmaz, amelyek diszulfidkötéssel kapcsolódnak egymáshoz, és egy további polipeptid J-láncot (1.31. ábra). Az ilyen formájú monomerek nehéz láncai nem tartalmaznak hidrofób farkat. A pentamernek 10 antigénkötő helye van, ezért hatékonyan felismeri és eltávolítja a szervezetbe először bekerülő antigént. Az IgM szekréciós formája az elsődleges immunválasz során a vérbe szekretált antitestek fő osztálya. Az IgM antigénhez való kötődése megváltoztatja az IgM konformációját, és indukálja kötődését a komplementrendszer első fehérjekomponenséhez (a komplementrendszer az antigén elpusztításában részt vevő fehérjék halmaza) és ennek a rendszernek az aktiválását. Ha az antigén egy mikroorganizmus felszínén helyezkedik el, a komplementrendszer a sejtmembrán integritásának megzavarását és a baktériumsejt halálát okozza.
Immunglobulinok G
Mennyiségileg az immunglobulinok ezen osztálya dominál a vérben (az összes Ig 75%-a). IgG - monomerek, a másodlagos immunválasz során a vérbe szekretált antitestek fő osztálya. Az IgG és a mikroorganizmusok felszíni antigénjei közötti kölcsönhatás után az antigén-antitest komplex képes megkötni és aktiválni a komplementrendszer fehérjéit, vagy kölcsönhatásba léphet a makrofágok és neutrofilek specifikus receptoraival. A fagocitákkal való kölcsönhatás vezet
 Rizs. 1.31. Az IgM szekréciós formájának felépítése
Rizs. 1.31. Az IgM szekréciós formájának felépítése
az antigén-antitest komplexek felszívódásához és sejtfagoszómákban való elpusztításához. Az IgG az egyetlen olyan antitestcsoport, amely képes áthatolni a placenta gáton, és méhen belüli védelmet nyújt a magzat számára a fertőzésekkel szemben.
Immunglobulinok A
A váladékban (tej, nyál, légúti és bélrendszeri váladék) található antitestek fő osztálya. Az IgA elsősorban dimer formában szekretálódik, ahol a monomerek egy további J-láncon keresztül kapcsolódnak egymáshoz (1.32. ábra).
Az IgA nem lép kölcsönhatásba a komplementrendszerrel és a fagocita sejtekkel, de a mikroorganizmusokhoz kötődve az antitestek megakadályozzák a hámsejtekhez való kötődésüket és a szervezetbe való behatolásukat.
Immunglobulinok E
Az E immunglobulinokat olyan monomerek képviselik, amelyekben a nehéz ε-láncok az M immunglobulinok μ-láncaihoz hasonlóan egy variábilis és négy konstans domént tartalmaznak. Az IgE a szekréció után kötődik azzal
 Rizs. 1.32. Az IgA szerkezete
Rizs. 1.32. Az IgA szerkezete
C-terminális régiók megfelelő receptorokkal a hízósejtek és bazofilek felszínén. Ennek eredményeként ezek a sejtek felületén antigének receptoraivá válnak (1.33. ábra).
 Rizs. 1.33. Az IgE és az antigén kölcsönhatása a hízósejt felszínén
Rizs. 1.33. Az IgE és az antigén kölcsönhatása a hízósejt felszínén
Miután az antigén az IgE megfelelő antigénkötő helyeihez kötődik, a sejtek jelet kapnak biológiailag aktív anyagok (hisztamin, szerotonin) kiválasztására, amelyek nagymértékben felelősek a gyulladásos reakció kialakulásáért és az allergiás reakciók megjelenéséért, mint pl. asztma, csalánkiütés, szénanátha.
Immunglobulinok D
A D immunglobulinok nagyon kis mennyiségben találhatók a szérumban, ezek monomerek. A nehéz δ láncoknak egy változó és három konstans doménje van. Az IgD-k a B-limfociták receptoraiként működnek, egyéb funkciók még nem ismertek. A specifikus antigének kölcsönhatása a B-limfociták (IgD) felszínén lévő receptorokkal ezeknek a jeleknek a sejtbe történő átviteléhez, illetve olyan mechanizmusok aktiválásához vezet, amelyek biztosítják az adott limfocita klón proliferációját.
TÉMAKÖR 1.7. A FEHÉRJÉK FIZIKAI ÉS KÉMIAI TULAJDONSÁGAI ÉS ELKÉPÍTÉSÜK MÓDSZEREI
1. Az egyes fehérjék fizikai és kémiai tulajdonságaikban különböznek:
Molekulák alakja;
Molekuláris tömeg;
A teljes töltés, amelynek nagysága az aminosavak anionos és kationos csoportjainak arányától függ;
A poláris és a nem poláris aminosav gyökök aránya a molekulák felületén;
Különféle denaturáló anyagokkal szembeni ellenállás foka.
2. A fehérje oldhatósága attól függ a fent felsorolt fehérjék tulajdonságairól, valamint a fehérjét feloldó táptalaj összetételéről (pH értékek, sóösszetétel, hőmérséklet, a fehérjével kölcsönhatásba lépő egyéb szerves anyagok jelenléte). A fehérjemolekulák töltési mennyisége az egyik olyan tényező, amely befolyásolja oldhatóságukat. Ha az izoelektromos pont töltése megszűnik, a fehérjék könnyebben aggregálódnak és kicsapódnak. Ez különösen jellemző a denaturált fehérjékre, amelyekben hidrofób aminosav gyökök jelennek meg a felszínen.
A fehérjemolekula felületén pozitív és negatív töltésű aminosavgyökök egyaránt találhatók. Ezeknek a csoportoknak a száma, így a fehérjék teljes töltése a táptalaj pH-jától függ, pl. a H+ - és OH - csoportok koncentrációinak aránya. Savas környezetben A H+ koncentráció növekedése a -COO - + H+ > -COOH karboxilcsoportok disszociációjának elnyomásához és a fehérjék negatív töltésének csökkenéséhez vezet. Lúgos környezetben az -NH 3 + + OH - - NH 2 + H 2 O aminocsoportok víz képződésével történő disszociációja során keletkező protonok OH-feleslegének megkötése a fehérjék pozitív töltésének csökkenéséhez vezet. . Azt a pH-értéket, amelynél a fehérje nettó nulla töltése van, nevezzük izoelektromos pont (IEP). Az IET-ben a pozitív és negatív töltésű csoportok száma azonos, i.e. a fehérje izoelektromos állapotban van.
3. Az egyes fehérjék szétválasztása. A test szerkezetének és működésének jellemzői a benne szintetizált fehérjék halmazától függenek. A fehérjék szerkezetének és tulajdonságainak tanulmányozása lehetetlen anélkül, hogy izolálnák őket a sejtből és megtisztítanák más fehérjéktől és szerves molekuláktól. Az egyes fehérjék izolálásának és tisztításának szakaszai:
Sejtpusztulás a vizsgált szövetet és homogenizátumot kapunk.
A homogenizátum szétválasztása frakciókra centrifugálással a kívánt fehérjét tartalmazó nukleáris, mitokondriális, citoszolos vagy más frakciót nyerünk.
Szelektív termikus denaturáció- fehérjeoldat rövid távú melegítése, amely során a denaturált fehérjeszennyeződések egy része eltávolítható (ha a fehérje viszonylag hőstabil).
Kisózni. A különböző fehérjék különböző sókoncentrációkkal válnak ki az oldatban. A sókoncentráció fokozatos növelésével számos különálló frakciót lehet előállítani, amelyek közül az egyikben túlnyomórészt izolált fehérje van. Az ammónium-szulfátot leggyakrabban fehérje frakcionálására használják. A legkevésbé oldódó fehérjék alacsony sókoncentráció esetén kicsapódnak.
Gélszűrés- Molekulák szitálásának módszere duzzadt Sephadex granulákon (a dextrán háromdimenziós poliszacharid láncai, amelyek pórusokkal rendelkeznek). Az a sebesség, amellyel a fehérjék áthaladnak a Sephadex-szel töltött oszlopon, a molekulatömegüktől függ: minél kisebb a fehérjemolekulák tömege, annál könnyebben hatolnak be a szemcsékbe, és minél hosszabb ideig maradnak ott, annál gyorsabban eluálódnak oszlop.
Ultracentrifugálás- olyan módszer, amely magában foglalja a fehérjéket egy centrifugacsőben az ultracentrifuga rotorjába helyezni. Amikor a rotor forog, a fehérjék ülepedési sebessége arányos molekulatömegükkel: a nehezebb fehérjék frakciói közelebb helyezkednek el a kémcső aljához, a könnyebbek - közelebb a felülethez.
Elektroforézis- a fehérjék elektromos térben való mozgási sebességének különbségén alapuló módszer. Ez az érték arányos a fehérjék töltésével. A fehérjék elektroforézisét papíron (ebben az esetben a fehérjék mozgásának sebessége csak a töltésükkel arányos) vagy egy bizonyos pórusméretű poliakrilamid gélben (a fehérjék mozgásának sebessége arányos a töltésükkel és molekulatömegükkel) végezzük. .
Ioncserélő kromatográfia- olyan frakcionálási módszer, amely ionizált fehérjecsoportok ioncserélő gyanta (oldhatatlan polimer anyagok) ellentétes töltésű csoportjaihoz való kötődésén alapul. A fehérje gyantához való kötődésének erőssége arányos a fehérje töltésével. Az ioncserélő polimerhez adszorbeált fehérjék a NaCl oldatok növekvő koncentrációjával lemoshatók; minél kisebb a fehérje töltése, annál kisebb a NaCl koncentrációja, amely a gyanta ionos csoportjaihoz kötött fehérje lemosásához szükséges.
Affinitáskromatográfia- a legspecifikusabb módszer az egyes fehérjék izolálására. Egy fehérje ligandumát kovalensen kötik egy inert polimerhez. Ha egy fehérjeoldatot egy polimerrel ellátott oszlopon vezetünk át, csak az adott ligandumra specifikus fehérje adszorbeálódik az oszlopon a fehérje ligandumhoz való komplementer kötődése miatt.
Dialízis- kis molekulatömegű vegyületek izolált fehérje oldatából való eltávolítására szolgáló módszer. A módszer azon alapul, hogy a fehérjék nem tudnak átjutni egy félig áteresztő membránon, ellentétben az alacsony molekulatömegű anyagokkal. A fehérjék kis molekulatömegű szennyeződésektől való megtisztítására szolgál, például a sók kisózását követően.
FELADATOK TANÁN KÍVÜLI MUNKÁHOZ
1. Töltse ki a táblázatot. 1.4.
1.4. táblázat. A rokon fehérjék - mioglobin és hemoglobin - szerkezetének és funkcióinak összehasonlító elemzése
a) emlékezzünk az Mb és Hb aktív központjának szerkezetére. Milyen szerepet játszanak a hidrofób aminosav gyökök e fehérjék aktív centrumainak kialakításában? Ismertesse az Mb és Hb aktív centrumának szerkezetét és a ligandumok kapcsolódási mechanizmusait! Milyen szerepet játszanak a His F 8 és His E 7 maradékok az Mv iHv aktív központjának működésében?
b) milyen új tulajdonságokkal rendelkezik a monomer mioglobinhoz képest a közeli rokon oligomer fehérje, a hemoglobin? Ismertesse a kooperatív változások szerepét a hemoglobin molekulában lévő protomerek konformációjában, a CO 2 és a protonkoncentráció hatását a hemoglobin oxigénaffinitására, valamint a 2,3-BPG szerepét a Hb funkció alloszterikus szabályozásában. .
2. Jellemezze a molekuláris chaperonokat, ügyelve szerkezetük és működésük kapcsolatára!
3. Milyen fehérjéket sorolunk családokba? Az immunglobulin család példáján azonosítsa a család fehérjéinek hasonló szerkezeti jellemzőit és kapcsolódó funkcióit.
4. Tisztított egyedi fehérjékre gyakran van szükség biokémiai és gyógyászati célokra. Ismertesse a fehérjék fiziko-kémiai tulajdonságain alapul az elválasztásukra és tisztításukra alkalmazott módszerek!
ÖNELLENŐRZÉSI FELADATOK
1. Válaszd ki a helyes válaszokat.
A hemoglobin funkciói:
A. O 2 szállítása a tüdőből a szövetbe B. H + szállítása a szövetekből a tüdőbe
B. A vér állandó pH-értékének fenntartása D. CO 2 szállítása a tüdőből a szövetekbe
D. CO 2 szállítása a szövetekből a tüdőbe
2. Válaszd ki a helyes válaszokat. Ligandα -Hb protomer: A. Heme
B. Oxigén
B. CO G. 2,3-BPG
D. β-protomer
3. Válaszd ki a helyes válaszokat.
Hemoglobin a mioglobinnal szemben:
A. Kvaterner szerkezetű
B. A másodlagos szerkezetet csak α-hélixek képviselik
B. Komplex fehérjékhez tartozik
D. Kölcsönhatásba lép egy allosztérikus ligandummal D. Kovalens kötéssel kötődik a hemhez
4. Válaszd ki a helyes válaszokat.
A Hb O2 iránti affinitása csökken:
A. Ha egy O 2 molekulát adunk hozzá B. Ha egy O 2 molekulát eltávolítunk
B. Amikor kölcsönhatásba lép a 2,3-BPG-vel
D. Ha protomerekhez kapcsolódnak H + D. Amikor a 2,3-BPG koncentrációja csökken
5. Mérkőzés.
A HB típusokat a következők jellemzik:
A. Dezoxi formában fibrilláris aggregátumokat képez B. Két α- és két δ-láncot tartalmaz
B. A Hb domináns formája a felnőtt eritrocitákban D. Hemet tartalmaz Fe+ 3-mal az aktív központban
D. Két α- és két γ-láncot tartalmaz 1. HbA 2.
6. Mérkőzés.
Hb ligandumok:
A. Hb-hez kötődik az allosztérikus központban
B. Nagyon nagy affinitása van a Hb aktív helyéhez
B. Csatlakozással növeli a Hb O 2 G iránti affinitását. A Fe+ 2 -t Fe+ 3 -dá oxidálja
D. Kovalens kötést képez a hisF8-cal
7. Válaszd ki a helyes válaszokat.
Kísérők:
A. A sejt minden részében jelenlévő fehérjék
B. A szintézis fokozódik stressz hatására
B. Részt vesz a denaturált fehérjék hidrolízisében
D. Részvétel a fehérjék natív konformációjának fenntartásában
D. Hozzon létre organellumokat, amelyekben a fehérjék konformációja képződik
8. Match. Immunglobulinok:
A. A szekréciós forma pentamer.
B. Ig osztály, amely áthatol a placenta gáton
B. Ig - hízósejt receptor
D. A hámsejtek váladékában jelen lévő Ig fő osztálya. D. B-limfocita receptor, melynek aktiválása biztosítja a sejtszaporodást
9. Válaszd ki a helyes válaszokat.
Immunglobulinok E:
A. Makrofágok termelik B. Nehéz ε láncokkal rendelkeznek.
B. T-limfociták membránjába ágyazva
D. Membrán antigén receptorként működnek hízósejteken és bazofileken
D. Felelős az allergiás reakciókért
10. Válaszd ki a helyes válaszokat.
A fehérjék elválasztásának módszere molekulatömegük különbségén alapul:
A. Gélszűrés
B. Ultracentrifugálás
B. Poliakrilamid gélelektroforézis D. Ioncserélő kromatográfia
D. Affinitáskromatográfia
11. Válaszd ki a megfelelő választ.
A fehérjék elválasztásának módszere a vízben való oldhatóságuk különbségén alapul:
A. Gélszűrés B. Kisózás
B. Ioncserélő kromatográfia D. Affinitáskromatográfia
D. Elektroforézis poliakrilamid gélben
AZ „ÖNELLENŐRZÉSI FELADATOKRA” VONATKOZÓ VÁLASZOK SZABVÁNYAI
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
ALAPVETŐ FELTÉTELEK ÉS FOGALMAK
1. Oligomer fehérjék, protomerek, fehérjék kvaterner szerkezete
2. Kooperatív változások a protomer konformációban
3. Bohr-effektus
4. Fehérjefunkciók alloszterikus szabályozása, alloszterikus centrum és alloszterikus effektor
5. Molekuláris chaperonok, hősokk fehérjék
6. Fehérjecsaládok (szerin proteázok, immunglobulinok)
7. IgM-, G-, E-, A-struktúra-függvény kapcsolat
8. Fehérjék össztöltése, fehérjék izoelektromos pontja
9. Elektroforézis
10. Kisózni
11. Gélszűrés
12. Ioncserélő kromatográfia
13. Ultracentrifugálás
14. Affinitáskromatográfia
15. Vérplazmafehérjék elektroforézise
FELADATOK A TANÉLETI MUNKÁHOZ
1. Hasonlítsa össze a hemoglobin (Hb) és a mioglobin (Mb) oxigéntel való telítési fokának a szövetekben fennálló parciális nyomásától való függését
 Rizs. 1.34. Telítettség függése Mv ésNHoxigént a parciális nyomásából
Rizs. 1.34. Telítettség függése Mv ésNHoxigént a parciális nyomásából
Kérjük, vegye figyelembe, hogy a fehérje oxigéntelítettségi görbéinek alakja eltérő: mioglobin esetében - hiperbola, hemoglobin esetében - szigma alakú.
1. Hasonlítsa össze az oxigén parciális nyomásának értékét, amelynél az Mb és a Hb 50%-kal telített O 2 -vel. Az alábbi fehérjék közül melyiknek van nagyobb affinitása az O 2 iránt?
2. Milyen szerkezeti jellemzők határozzák meg az Mb O 2 iránti nagy affinitását?
3. A HB milyen szerkezeti jellemzői teszik lehetővé, hogy a nyugalmi szövetek kapillárisaiban O2-t szabadítson fel (viszonylag magas O2 parciális nyomás mellett), és ezt a felszabadulást erőteljesen növelje a dolgozó izmokban? Az oligomer fehérjék milyen tulajdonsága biztosítja ezt a hatást?
4. Számolja ki, mekkora O 2 (%-ban) oxigéntartalmú hemoglobint ad a pihenő és dolgozó izmoknak?
5. vonjon le következtetéseket a fehérje szerkezete és működése közötti összefüggésre.
2. A hemoglobin által a kapillárisokban felszabaduló oxigén mennyisége a szövetekben zajló katabolikus folyamatok intenzitásától függ (Bohr-effektus). Hogyan szabályozzák a szöveti anyagcsere változásai a Hb O2 iránti affinitását? A CO 2 és a H+ hatása a Hb O 2 iránti affinitására
 1. írja le a Bohr-effektust.
1. írja le a Bohr-effektust.
2. milyen irányban halad a diagramon látható folyamat:
a) a tüdő kapillárisaiban;
b) szöveti kapillárisokban?
3. Mi a Bohr-effektus élettani jelentősége?
4. Miért változtatja meg a Hb és a H+ kölcsönhatása a hemtől távoli helyeken a fehérje O 2 iránti affinitását?
3. A Hb O2-hoz való affinitása ligandjának, a 2,3-biszfoszfoglicerátnak a koncentrációjától függ, amely a Hb O2 iránti affinitásának alloszterikus szabályozója. Miért befolyásolja a ligandum kölcsönhatás az aktív helytől távoli helyen a fehérje működését? Hogyan szabályozza a 2,3-BPG a Hb O2 iránti affinitását? A probléma megoldásához válaszoljon a következő kérdésekre:
1. hol és miből szintetizálják a 2,3-biszfoszfoglicerátot (2,3-BPG)? Írd fel a képletét, jelöld meg ennek a molekulának a töltését!
2. A hemoglobin melyik formájával (oxi vagy dezoxi) lép kölcsönhatásba a BPG és miért? A Hb molekula melyik részében jön létre a kölcsönhatás?
3. milyen irányba halad az ábrán látható folyamat?
a) szöveti kapillárisokban;
b) a tüdő kapillárisaiban?
4. ahol a komplex koncentrációja nagyobb legyen
Nv-2,3-BFG:
a) nyugalmi izmok hajszálereiben,
b) a dolgozó izmok kapillárisaiban (azonos BPG-koncentráció mellett az eritrocitákban)?
5. Hogyan változik a HB oxigén iránti affinitása, amikor az ember alkalmazkodik a magas tengerszint feletti körülményekhez, ha megnő a BPG koncentrációja az eritrocitákban? Mi ennek a jelenségnek a fiziológiai jelentősége?
4. A 2,3-BPG pusztulása a tartósított vér tárolása során rontja a HB funkcióit. Hogyan változik a HB O 2 iránti affinitása a tartósított vérben, ha a 2,3-BPG koncentrációja az eritrocitákban 8-ról 0,5 mmol/l-re csökkenhet. Lehetséges-e ilyen vér transzfúziót adni súlyos betegeknek, ha a 2,3-BPG koncentrációja legkorábban három nap múlva áll helyre? Visszaállítható-e a vörösvértestek funkciója 2,3-BPG vérbe adásával?
5. Emlékezzen a legegyszerűbb immunglobulin molekulák szerkezetére. Milyen szerepet játszanak az immunglobulinok az immunrendszerben? Miért nevezik az Ig-eket gyakran bivalenseknek? Hogyan kapcsolódik az Ig-ek szerkezete funkciójukhoz? (Írja le az immunglobulinok egy osztályának példáján.)
A fehérjék fizikai-kémiai tulajdonságai és elválasztásuk módszerei.
6. Hogyan befolyásolja a fehérje nettó töltése az oldhatóságát?
a) határozzuk meg a peptid teljes töltését pH 7-nél
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) hogyan változik ennek a peptidnek a töltése pH >7, pH mellett<7, рН <<7?
c) mi a fehérje izoelektromos pontja (IEP), és milyen környezetben található?
Ennek a peptidnek az IET-je?
d) milyen pH-értéknél figyelhető meg ennek a peptidnek a legalacsonyabb oldhatósága.
7. Miért „alvadékosodik meg” a friss tejjel ellentétben a savanyú tej forraláskor (azaz a tejfehérje kazein kicsapódik)? A friss tejben a kazein molekulák negatív töltéssel rendelkeznek.
8. Az egyes fehérjék elkülönítésére gélszűrést alkalmaznak. A 160 000, 80 000 és 60 000 molekulatömegű A, B, C fehérjéket tartalmazó keveréket gélszűréssel elemeztük (1.35. ábra). A megduzzadt gélszemcsék áteresztők a 70 000-nél kisebb molekulatömegű fehérjék számára. Milyen elvek alapján áll ez az elválasztási módszer? Melyik gráf tükrözi helyesen a frakcionálás eredményeit? Adja meg, hogy az A, B és C fehérjék milyen sorrendben szabadulnak fel az oszlopról.
 Rizs. 1.35. Gélszűrés használata a fehérjék szétválasztására
Rizs. 1.35. Gélszűrés használata a fehérjék szétválasztására
9. ábrán. 1.36, A egy diagramot mutat az elektroforézisről papíron egy egészséges ember vérszérumfehérjéiről. Az ezzel a módszerrel kapott fehérjefrakciók relatív mennyiségei: albuminok 54-58%, α1-globulinok 6-7%, α2-globulinok 8-9%, β-globulinok 13%, γ-globulinok 11-12%.
 Rizs. 1.36 Elektroforézis papíron egészséges ember (A) és beteg (B) vérplazmafehérjéiről
Rizs. 1.36 Elektroforézis papíron egészséges ember (A) és beteg (B) vérplazmafehérjéiről
I - y-globulinok; II - β-globulinok; III -α 2-globulin; IV -α 2-globulin; V - albuminok
Sok betegséget kísérnek a szérumfehérjék összetételének mennyiségi változásai (dysproteinémia). E változások természetét a diagnózis felállításakor, valamint a betegség súlyosságának és stádiumának értékelése során figyelembe veszik.
Mókusok- α-aminosav-maradékokból álló, nagy molekulatömegű szerves vegyületek.
BAN BEN fehérje összetétel magában foglalja a szenet, hidrogént, nitrogént, oxigént, ként. Egyes fehérjék komplexeket képeznek más foszfort, vasat, cinket és rezet tartalmazó molekulákkal.
A fehérjék nagy molekulatömegűek: tojásalbumin - 36 000, hemoglobin - 152 000, miozin - 500 000 Összehasonlításképpen: az alkohol molekulatömege 46, az ecetsav - 60, a benzol - 78.
A fehérjék aminosav összetétele
Mókusok- nem periodikus polimerek, amelyek monomerjei α-aminosavak. Általában 20 fajta α-aminosavat neveznek fehérjemonomernek, bár ezek közül több mint 170 megtalálható a sejtekben és a szövetekben.
Attól függően, hogy az aminosavak szintetizálhatók-e az emberek és más állatok testében, megkülönböztetik őket: nem esszenciális aminosavak- szintetizálható; esszenciális aminosavak- nem szintetizálható. Az esszenciális aminosavakat táplálékkal kell bevinni a szervezetbe. A növények minden típusú aminosavat szintetizálnak.
Az aminosav összetételtől függően fehérjék: teljes- tartalmazza az aminosavak teljes készletét; hibás- néhány aminosav hiányzik az összetételükből. Ha a fehérjék csak aminosavakból állnak, akkor ún egyszerű. Ha a fehérjék az aminosavak mellett nem aminosav komponenst (protéziscsoportot) is tartalmaznak, akkor ún. összetett. A protéziscsoportot fémek (fémproteinek), szénhidrátok (glikoproteinek), lipidek (lipoproteinek), nukleinsavak (nukleoproteinek) képviselhetik.
Minden aminosavak tartalmaznak: 1) karboxilcsoport (-COOH), 2) aminocsoport (-NH 2), 3) gyök vagy R-csoport (a molekula többi része). A gyök szerkezete eltérő a különböző típusú aminosavak esetében. Az aminosavak összetételében szereplő aminocsoportok és karboxilcsoportok számától függően megkülönböztetik őket: semleges aminosavak egy karboxilcsoporttal és egy aminocsoporttal rendelkezik; bázikus aminosavak egynél több aminocsoporttal rendelkezik; savas aminosavak egynél több karboxilcsoportot tartalmaz.
Az aminosavak azok amfoter vegyületek, mivel oldatban savként és bázisként is működhetnek. Vizes oldatokban az aminosavak különböző ionos formákban léteznek.
Peptid kötés
Peptidek- peptidkötésekkel összekapcsolt aminosav-maradékokból álló szerves anyagok.
A peptidek képződése az aminosavak kondenzációs reakciójának eredményeként megy végbe. Amikor az egyik aminosav aminocsoportja kölcsönhatásba lép egy másik aminosav karboxilcsoportjával, kovalens nitrogén-szén kötés jön létre közöttük, amit ún. peptid. A peptidben található aminosavak számától függően vannak dipeptidek, tripeptidek, tetrapeptidek stb. A peptidkötés kialakulása sokszor megismételhető. Ez a formációhoz vezet polipeptidek. A peptid egyik végén egy szabad aminocsoport (úgynevezett N-terminális), a másik végén pedig egy szabad karboxilcsoport (úgynevezett C-terminális) található.

A fehérjemolekulák térbeli szerveződése
A fehérjék bizonyos specifikus funkcióinak ellátása molekuláik térbeli konfigurációjától függ, emellett a sejt számára energetikailag kedvezőtlen, ha a fehérjéket kibontott formában, lánc formájában tartja, ezért a polipeptid láncok feltekerednek, s így feltekernek. bizonyos háromdimenziós szerkezet vagy konformáció. 4 szint van fehérjék térbeli szerveződése.
Elsődleges fehérje szerkezet- a fehérjemolekulát alkotó polipeptidlánc aminosavak elrendeződésének sorrendje. Az aminosavak közötti kötés peptidkötés.

Ha egy fehérjemolekula csak 10 aminosavból áll, akkor a fehérjemolekulák elméletileg lehetséges változatainak száma, amelyek az aminosavak váltakozási sorrendjében különböznek egymástól, 10 20. 20 aminosav birtokában még változatosabb kombinációkat készíthet belőlük. Körülbelül tízezer különböző fehérjét találtak az emberi szervezetben, amelyek mind egymástól, mind más szervezetek fehérjéitől különböznek.
Ez a fehérjemolekula elsődleges szerkezete, amely meghatározza a fehérjemolekulák tulajdonságait és térbeli konfigurációját. Ha csak egy aminosavat helyettesítünk egy másikkal a polipeptidláncban, az a fehérje tulajdonságainak és funkcióinak megváltozásához vezet. Például a hatodik glutamin aminosav helyettesítése valinnal a hemoglobin β-alegységében ahhoz a tényhez vezet, hogy a hemoglobin molekula egésze nem tudja ellátni fő funkcióját - oxigénszállítást; Ilyen esetekben az emberben sarlósejtes vérszegénységnek nevezett betegség alakul ki.
Másodlagos szerkezet- a polipeptidlánc spirálba rendezett hajtogatása (úgy néz ki, mint egy kiterjesztett rugó). A hélix fordulatait a karboxilcsoportok és az aminocsoportok között létrejövő hidrogénkötések erősítik. A hidrogénkötések kialakításában szinte minden CO és NH csoport részt vesz. Gyengébbek, mint a peptidek, de sokszor megismételve stabilitást és merevséget kölcsönöznek ennek a konfigurációnak. A másodlagos szerkezet szintjén fehérjék találhatók: fibroin (selyem, pókháló), keratin (haj, köröm), kollagén (inak).



Harmadlagos szerkezet- kémiai kötések (hidrogén, ionos, diszulfid) képződéséből és az aminosavmaradékok gyökei közötti hidrofób kölcsönhatások létrejöttéből adódó polipeptid láncok gömbökbe tömörítése. A harmadlagos szerkezet kialakításában a fő szerepet a hidrofil-hidrofób kölcsönhatások játsszák. A vizes oldatokban a hidrofób gyökök hajlamosak elbújni a víz elől, csoportosulva a gömbölyű belsejében, míg a hidrofil gyökök a hidratáció (vízdipólusokkal való kölcsönhatás) következtében a molekula felületén jelennek meg. Egyes fehérjékben a harmadlagos szerkezetet két ciszteinmaradék kénatomjai között kialakuló diszulfid kovalens kötések stabilizálják. A harmadlagos szerkezeti szinten enzimek, antitestek és bizonyos hormonok találhatók.

Negyedidős szerkezet olyan komplex fehérjékre jellemző, amelyek molekuláit két vagy több gömböcske alkotja. Az alegységeket ionos, hidrofób és elektrosztatikus kölcsönhatások tartják a molekulában. Néha a kvaterner szerkezet kialakulása során diszulfid kötések lépnek fel az alegységek között. A legtöbbet vizsgált kvaterner szerkezetű fehérje az hemoglobin. Két α-alegység (141 aminosav) és két β-alegység (146 aminosav) alkotja. Mindegyik alegységhez vasat tartalmazó hem molekula kapcsolódik.
Ha valamilyen okból a fehérjék térbeli konformációja eltér a normálistól, a fehérje nem tudja ellátni funkcióit. Például a „bolondmarha-kór” (spongiform encephalopathia) oka a prionok, az idegsejtek felszíni fehérjéinek abnormális konformációja.
A fehérjék tulajdonságai
A fehérjemolekula aminosav-összetétele és szerkezete határozza meg tulajdonságait. A fehérjék bázikus és savas tulajdonságokat kombinálnak, amelyeket az aminosav gyökök határoznak meg: minél több savas aminosav van egy fehérjében, annál kifejezettebb a savas tulajdonságai. Meg van határozva az adományozás és a H + hozzáadásának képessége a fehérjék pufferelő tulajdonságai; Az egyik legerősebb puffer a vörösvérsejtekben lévő hemoglobin, amely a vér pH-ját állandó szinten tartja. Vannak oldható fehérjék (fibrinogén), és vannak oldhatatlan fehérjék, amelyek mechanikai funkciókat látnak el (fibroin, keratin, kollagén). Vannak olyan fehérjék, amelyek kémiailag aktívak (enzimek), vannak kémiailag inaktívak, amelyek ellenállnak a különböző környezeti feltételeknek, és amelyek rendkívül instabilak.
Külső tényezők (hő, ultraibolya sugárzás, nehézfémek és sóik, pH-változások, sugárzás, kiszáradás)
zavart okozhat a fehérjemolekula szerkezeti szerveződésében. Az adott fehérjemolekulában rejlő háromdimenziós konformáció elvesztésének folyamatát ún denaturáció. A denaturáció oka egy bizonyos fehérjeszerkezetet stabilizáló kötések felszakadása. Kezdetben a leggyengébb kötelékek szakadnak meg, és a feltételek szigorodásával még erősebbek is megszakadnak. Ezért először a kvaterner, majd a harmadlagos és másodlagos szerkezetek vesznek el. A térbeli konfiguráció változása a fehérje tulajdonságainak megváltozásához vezet, és ennek eredményeként lehetetlenné teszi a fehérje számára az eredendő biológiai funkcióinak ellátását. Ha a denaturációt nem kíséri az elsődleges szerkezet pusztulása, akkor lehet megfordítható, ebben az esetben megtörténik a fehérjére jellemző konformáció öngyógyítása. Például a membránreceptor fehérjék ilyen denaturáción mennek keresztül. A fehérjeszerkezet denaturáció utáni helyreállításának folyamatát ún renaturáció. Ha a fehérje térbeli konfigurációjának helyreállítása lehetetlen, akkor denaturációt nevezünk visszafordíthatatlan.
A fehérjék funkciói
| Funkció | Példák és magyarázatok |
|---|---|
| Építkezés | A fehérjék részt vesznek a sejtes és extracelluláris struktúrák kialakításában: a sejtmembránok (lipoproteinek, glikoproteinek), a haj (keratin), az inak (kollagén) stb. részei. |
| Szállítás | A hemoglobin vérfehérje oxigént köt, és azt a tüdőből minden szövetbe és szervbe szállítja, és onnan szén-dioxidot szállít a tüdőbe; A sejtmembránok összetétele speciális fehérjéket tartalmaz, amelyek biztosítják bizonyos anyagok és ionok aktív és szigorúan szelektív átvitelét a sejtből a külső környezetbe és vissza. |
| Szabályozó | A fehérjehormonok részt vesznek az anyagcsere folyamatok szabályozásában. Például az inzulin hormon szabályozza a vércukorszintet, elősegíti a glikogén szintézist, és fokozza a zsírok képződését a szénhidrátokból. |
| Védő | Az idegen fehérjék vagy mikroorganizmusok (antigének) szervezetbe való behatolására válaszul speciális fehérjék képződnek - antitestek, amelyek meg tudják kötni és semlegesíteni tudják őket. A fibrinogénből képződő fibrin segít megállítani a vérzést. |
| Motor | Az aktin és a miozin kontraktilis fehérjék biztosítják az izomösszehúzódást többsejtű állatokban. |
| Jel | A sejt felszíni membránjába olyan fehérjemolekulák vannak beépítve, amelyek a környezeti tényezők hatására képesek harmadlagos szerkezetük megváltoztatására, így a külső környezet jeleit fogadva, parancsokat továbbítva a sejtnek. |
| Tárolás | Az állatok testében a fehérjék általában nem raktározódnak, kivéve a tojásalbumint és a tejkazeint. De a fehérjéknek köszönhetően bizonyos anyagok raktározódhatnak a szervezetben, például a hemoglobin lebontása során a vas nem távozik a szervezetből, hanem elraktározódik, komplexet képezve a ferritinnel. |
| Energia | Amikor 1 g fehérje végtermékekre bomlik, 17,6 kJ szabadul fel. Először a fehérjék aminosavakra, majd végtermékekre - vízre, szén-dioxidra és ammóniára - bomlanak. A fehérjéket azonban csak akkor használják energiaforrásként, ha más forrásokat (szénhidrátokat és zsírokat) elhasználnak. |
| Katalitikus | A fehérjék egyik legfontosabb funkciója. A fehérjék biztosítják - enzimek, amelyek felgyorsítják a sejtekben előforduló biokémiai reakciókat. Például a ribulóz-bifoszfát-karboxiláz katalizálja a CO 2 rögzítését a fotoszintézis során. |
Enzimek
Enzimek, vagy enzimek, a fehérjék egy speciális osztálya, amelyek biológiai katalizátorok. Az enzimeknek köszönhetően a biokémiai reakciók óriási sebességgel mennek végbe. Az enzimes reakciók sebessége több tízezerszer (és néha milliókkal) nagyobb, mint a szervetlen katalizátorok részvételével lezajló reakciók sebessége. Az anyagot, amelyre az enzim hat, nevezzük szubsztrát.
Az enzimek globuláris fehérjék, szerkezeti jellemzők Az enzimek két csoportra oszthatók: egyszerű és összetett. Egyszerű enzimek egyszerű fehérjék, pl. csak aminosavakból áll. Komplex enzimek komplex fehérjék, pl. A fehérje részen kívül tartalmaznak egy nem fehérje jellegű csoportot is - kofaktor. Egyes enzimek vitaminokat használnak kofaktorként. Az enzimmolekula egy speciális részt tartalmaz, az úgynevezett aktív központot. Aktív központ- az enzim egy kis része (3-12 aminosav), ahol a szubsztrát vagy szubsztrátok megkötése enzim-szubsztrát komplexet képez. A reakció befejeződése után az enzim-szubsztrát komplex enzimre és reakciótermék(ek)re bomlik. Egyes enzimek (az aktívak kivételével) allosztérikus központok- olyan területek, amelyekhez enzimsebesség-szabályozók vannak csatlakoztatva ( alloszterikus enzimek).

Az enzimatikus katalízis reakcióit: 1) nagy hatékonyság, 2) szigorú szelektivitás és hatásirány, 3) szubsztrátspecifitás, 4) finom és precíz szabályozás jellemzi. Az enzimatikus katalízis reakciók szubsztrát- és reakcióspecifitását E. Fischer (1890) és D. Koshland (1959) hipotézise magyarázza.
E. Fischer (billentyűzár hipotézis) azt javasolta, hogy az enzim aktív centrumának és a szubsztrátnak a térbeli konfigurációinak pontosan meg kell felelniük egymásnak. A szubsztrátot a „kulcshoz”, az enzimet a „zárhoz” hasonlítják.
D. Koshland (kéz-kesztyű hipotézis) azt javasolta, hogy a szubsztrát szerkezete és az enzim aktív centruma közötti térbeli megfelelés csak az egymással való kölcsönhatás pillanatában jön létre. Ezt a hipotézist más néven indukált megfelelési hipotézis.
Az enzimreakciók sebessége függ: 1) hőmérséklettől, 2) enzimkoncentrációtól, 3) szubsztrátkoncentrációtól, 4) pH-tól. Hangsúlyozni kell, hogy mivel az enzimek fehérjék, aktivitásuk fiziológiailag normális körülmények között a legmagasabb.
A legtöbb enzim csak 0 és 40°C közötti hőmérsékleten tud működni. Ezen határokon belül a reakciósebesség körülbelül 2-szeresére nő minden 10 °C-os hőmérséklet-emelkedéssel. 40 °C feletti hőmérsékleten a fehérje denaturálódik, és az enzimaktivitás csökken. Fagyponthoz közeli hőmérsékleten az enzimek inaktiválódnak.
A szubsztrát mennyiségének növekedésével az enzimatikus reakció sebessége addig növekszik, amíg a szubsztrátmolekulák száma el nem éri az enzimmolekulák számát. A szubsztrát mennyiségének további növelésével a sebesség nem növekszik, mivel az enzim aktív központjai telítettek. Az enzimkoncentráció növekedése megnövekedett katalitikus aktivitáshoz vezet, mivel egységnyi idő alatt nagyobb számú szubsztrátmolekula átalakul.

Mindegyik enzimhez van egy optimális pH-érték, amelynél maximális aktivitást mutat (pepszin - 2,0, nyálamiláz - 6,8, hasnyálmirigy-lipáz - 9,0). Magasabb vagy alacsonyabb pH-értékeknél az enzimaktivitás csökken. A pH hirtelen változásával az enzim denaturálódik.
Az alloszterikus enzimek sebességét az alloszterikus központokhoz kapcsolódó anyagok szabályozzák. Ha ezek az anyagok felgyorsítják a reakciót, akkor ún aktivátorok, ha lelassulnak - inhibitorok.
Az enzimek osztályozása
Az általuk katalizált kémiai átalakulások típusa alapján az enzimeket 6 osztályba sorolják:
- oxireduktázok(hidrogén-, oxigén- vagy elektronatomok átvitele egyik anyagból a másikba - dehidrogenáz),
- transzferázok(metil-, acil-, foszfát- vagy aminocsoport átvitele egyik anyagból a másikba - transzamináz),
- hidrolázok(hidrolízis reakciók, amelyek során két termék képződik a szubsztrátból - amiláz, lipáz),
- lyázok(nem hidrolitikus addíció a szubsztrátumhoz vagy atomcsoport leválasztása róla, ilyenkor a C-C, C-N, C-O, C-S kötések felszakadhatnak - dekarboxiláz),
- izomerázok(intramolekuláris átrendeződés - izomeráz),
- ligázok(két molekula összekapcsolódása C-C, C-N, C-O, C-S kötések kialakulásának eredményeként - szintetáz).
Az osztályokat alosztályokra és alosztályokra osztják fel. A jelenlegi nemzetközi osztályozásban minden enzimnek saját kódja van, amely négy, pontokkal elválasztott számból áll. Az első szám az osztály, a második az alosztály, a harmadik az alosztály, a negyedik az enzim sorozatszáma ebben az alosztályban, például az argináz kód 3.5.3.1.
Menj előadások 2. sz"A szénhidrátok és lipidek szerkezete és funkciói"
Menj előadások 4. sz"Az ATP nukleinsavak szerkezete és funkciói"