19. sajandi esimesel poolel. paljud keemikud, nende hulgas eelkõige J. von Liebig, jõudsid järk-järgult järeldusele, et valgud moodustavad lämmastikuühendite eriklassi. Nimi "valgud" (kreeka keelest.
protod esimene) pakkus 1840. aastal välja Hollandi keemik G. Mulder. FÜÜSIKALISED OMADUSED Valgud on tahkes olekus valged, kuid lahuses värvitud, välja arvatud juhul, kui neil on mingi kromofoor (värviline) rühm, näiteks hemoglobiin. Erinevate valkude lahustuvus vees on väga erinev. See muutub ka sõltuvalt pH-st ja soolade kontsentratsioonist lahuses, seega on võimalik valida tingimused, mille korral üks valk sadestub selektiivselt teiste valkude juuresolekul. Seda "väljasoolamise" meetodit kasutatakse laialdaselt valkude eraldamiseks ja puhastamiseks. Puhastatud valk sadestub sageli lahusest välja kristallidena.Võrreldes teiste ühenditega on valkude molekulmass väga suur, ulatudes mitmest tuhandest kuni mitme miljoni daltonini. Seetõttu settivad ultratsentrifuugimise ajal valgud erineva kiirusega. Positiivse ja negatiivse laenguga rühmade esinemise tõttu valgumolekulides liiguvad need erineva kiirusega ja elektriväljas. See on elektroforeesi alus, meetod, mida kasutatakse üksikute valkude eraldamiseks keerukatest segudest. Valke puhastatakse ka kromatograafiaga.
KEEMILISED OMADUSED Struktuur. Valgud on polümeerid, st. molekulid, mis on ehitatud nagu ahelad korduvatest monomeerüksustest või subühikutest, mille rolli nad mängivad a -aminohapped. Aminohapete üldvalem kus R vesinikuaatom või mõni orgaaniline rühm.Valgu molekul (polüpeptiidahel) võib koosneda ainult suhteliselt väikesest arvust aminohapetest või mitmest tuhandest monomeerühikust. Aminohapete kombineerimine ahelas on võimalik, kuna igaühel neist on kaks erinevat keemilist rühma: aluseliste omadustega aminorühm,
NH2 ja happeline karboksüülrühm COOH. Mõlemad rühmad on seotud a -süsiniku aatom. Ühe aminohappe karboksüülrühm võib moodustada amiid- (peptiid-) sideme teise aminohappe aminorühmaga: Pärast seda, kui kaks aminohapet on sel viisil ühendatud, saab ahelat pikendada, lisades teisele aminohappele kolmanda jne. Nagu ülaltoodud võrrandist näha, vabaneb peptiidsideme moodustumisel vee molekul. Hapete, leeliste või proteolüütiliste ensüümide juuresolekul kulgeb reaktsioon vastupidises suunas: polüpeptiidahel jagatakse vee lisamisega aminohapeteks. Seda reaktsiooni nimetatakse hüdrolüüsiks. Hüdrolüüs toimub spontaanselt ja aminohapete ühendamiseks polüpeptiidahelaks on vaja energiat.
Pärast seda, kui kaks aminohapet on sel viisil ühendatud, saab ahelat pikendada, lisades teisele aminohappele kolmanda jne. Nagu ülaltoodud võrrandist näha, vabaneb peptiidsideme moodustumisel vee molekul. Hapete, leeliste või proteolüütiliste ensüümide juuresolekul kulgeb reaktsioon vastupidises suunas: polüpeptiidahel jagatakse vee lisamisega aminohapeteks. Seda reaktsiooni nimetatakse hüdrolüüsiks. Hüdrolüüs toimub spontaanselt ja aminohapete ühendamiseks polüpeptiidahelaks on vaja energiat. Karboksüülrühm ja amiidrühm (või sarnane imiidrühm aminohappe proliini puhul) on olemas kõigis aminohapetes, kuid aminohapete erinevused on määratud rühma või kõrvalahela olemusega. mis on ülalpool kirjaga tähistatud
R . Külgahela rolli võib täita üks vesinikuaatom, nagu aminohappes glütsiin, või mõni mahukas rühm, nagu histidiinis ja trüptofaanis. Mõned külgahelad on keemiliselt inertsed, teised aga märkimisväärselt reaktiivsed.Sünteesida saab palju tuhandeid erinevaid aminohappeid ja looduses esineb palju erinevaid aminohappeid, kuid valgusünteesiks kasutatakse ainult 20 tüüpi aminohappeid: alaniin, arginiin, asparagiin, asparagiinhape, valiin, histidiin, glütsiin, glutamiin, glutamiin hape, isoleutsiin, leutsiin, lüsiin, metioniin, proliin, seriin, türosiin, treoniin, trüptofaan, fenüülalaniin ja tsüsteiin (valkudes võib tsüsteiin esineda dimeerina
tsüstiin). Tõsi, mõned valgud sisaldavad lisaks regulaarselt esinevale kahekümnele ka teisi aminohappeid, kuid need tekivad ühe kahekümnest loetletud modifikatsiooni tulemusena pärast selle lisamist valgu hulka.Optiline aktiivsus. Kõik aminohapped, välja arvatud glütsiin, on a -süsiniku aatomiga on seotud neli erinevat rühma. Geomeetria seisukohalt saab nelja erinevat rühma kinnitada kahel viisil ja vastavalt sellele on kaks võimalikku konfiguratsiooni ehk kaks isomeeri, mis on omavahel seotud nagu objekt on selle peegelpildiga, s.t. nagu vasak käsi paremale. Ühte konfiguratsiooni nimetatakse vasakpoolseks või vasakukäeliseks ( L ) ja teine parempoolne või paremale pöörav ( D ), kuna kaks sellist isomeeri erinevad polariseeritud valguse tasandi pöörlemissuuna poolest. Leidub ainult valkudes L -aminohapped (erandiks on glütsiin; seda saab esitada ainult ühel kujul, kuna selle neljast rühmast kaks on samad) ja kõik need on optiliselt aktiivsed (kuna on ainult üks isomeer). D -aminohapped on looduses haruldased; neid leidub mõnes antibiootikumis ja bakterite rakuseinas.Aminohapete järjestus. Polüpeptiidahelas ei ole aminohapped paigutatud juhuslikult, vaid kindlas kindlas järjekorras ning just see järjestus määrab valgu funktsioonid ja omadused. Varieerides 20 tüüpi aminohapete järjestust, saate luua tohutul hulgal erinevaid valke, nii nagu saate luua palju erinevaid tekste tähestiku tähtedest.Varem kestis valgu aminohappejärjestuse määramine sageli mitu aastat. Otsene määramine on endiselt üsna töömahukas ülesanne, kuigi on loodud seadmed, mis võimaldavad seda automaatselt teostada. Tavaliselt on lihtsam määrata vastava geeni nukleotiidjärjestus ja tuletada sellest valgu aminohappejärjestus. Tänaseks on paljude sadade valkude aminohappejärjestused juba kindlaks määratud. Dešifreeritud valkude funktsioonid on tavaliselt teada ja see aitab ette kujutada sarnaste valkude võimalikke funktsioone, mis tekivad näiteks pahaloomulistes kasvajates.
Komplekssed valgud. Ainult aminohapetest koosnevaid valke nimetatakse lihtsateks. Sageli on aga polüpeptiidahela külge kinnitatud metalliaatom või mõni keemiline ühend, mis ei ole aminohape. Selliseid valke nimetatakse kompleksseteks. Näiteks hemoglobiin: see sisaldab raudporfüriini, mis määrab selle punase värvuse ja võimaldab tal toimida hapnikukandjana.Enamike keerukamate valkude nimetused näitavad seotud rühmade olemust: glükoproteiinid sisaldavad suhkruid, lipoproteiinid sisaldavad rasvu. Kui ensüümi katalüütiline aktiivsus sõltub seotud rühmast, siis nimetatakse seda proteesrühmaks. Sageli mängib vitamiin proteesirühma rolli või on selle osa. Näiteks A-vitamiin, mis on kinnitunud ühe võrkkesta valgu külge, määrab selle valgustundlikkuse.
Tertsiaarne struktuur. Tähtis pole mitte niivõrd valgu enda aminohappejärjestus (esmane struktuur), vaid viis, kuidas see ruumis paikneb. Kogu polüpeptiidahela pikkuses moodustavad vesinikioonid korrapäraseid vesiniksidemeid, mis annavad sellele spiraali või kihi kuju (sekundaarne struktuur). Selliste spiraalide ja kihtide kombinatsioonist tekib järgmise järjestuse kompaktne vorm: valgu tertsiaarne struktuur. Ahela monomeerüksusi hoidvate sidemete ümber on võimalikud väikeste nurkade all pöörlemised. Seetõttu on puhtalt geomeetrilisest vaatenurgast mis tahes polüpeptiidahela võimalike konfiguratsioonide arv lõpmatult suur. Tegelikkuses eksisteerib iga valk tavaliselt ainult ühes konfiguratsioonis, mille määrab tema aminohappejärjestus. See struktuur ei ole jäik, see on justkui « hingab” kõigub teatud keskmise konfiguratsiooni ümber. Ahel on volditud konfiguratsiooni, milles vaba energia (võime toota tööd) on minimaalne, nii nagu vabastatud vedru surub kokku ainult minimaalsele vabaenergiale vastavasse olekusse. Sageli on üks ahela osa teisega jäigalt seotud disulfiidiga ( SS) sidemed kahe tsüsteiinijäägi vahel. Osaliselt seetõttu on tsüsteiinil aminohapete hulgas eriti oluline roll.Valkude struktuuri keerukus on nii suur, et valgu tertsiaarset struktuuri pole veel võimalik välja arvutada, isegi kui selle aminohappejärjestus on teada. Kuid kui on võimalik saada valgukristalle, saab selle tertsiaarset struktuuri määrata röntgendifraktsiooniga.
Struktuursetes, kokkutõmbumisvalkudes ja mõnedes teistes valkudes on ahelad piklikud ja mitmed läheduses asuvad kergelt volditud ahelad moodustavad fibrillid; fibrillid omakorda murduvad suuremateks kiududeks. Enamus lahuses olevaid valke on aga keraja kujuga: ahelad on kerakujuliselt keritud nagu lõng keras. Selle konfiguratsiooniga on vaba energia minimaalne, kuna hüdrofoobsed ("vett tõrjuvad") aminohapped on peidus kera sees ja hüdrofiilsed ("vett tõmbavad") aminohapped on selle pinnal.
Paljud valgud on mitme polüpeptiidahela kompleksid. Seda struktuuri nimetatakse valgu kvaternaarseks struktuuriks. Näiteks hemoglobiini molekul koosneb neljast alaühikust, millest igaüks on globulaarne valk.
Struktuurvalgud moodustavad oma lineaarse konfiguratsiooni tõttu väga kõrge tõmbetugevusega kiude, samal ajal kui globulaarne konfiguratsioon võimaldab valkudel astuda spetsiifilistesse interaktsioonidesse teiste ühenditega. Kui ahelad on õigesti paigutatud, ilmuvad kera pinnale teatud kujuga õõnsused, milles asuvad reaktiivsed keemilised rühmad. Kui antud valk on ensüüm, siis sellisesse õõnsusse siseneb mingi aine teine, tavaliselt väiksem molekul, täpselt nagu võti lukku; sel juhul muutub õõnsuses paiknevate keemiliste rühmade mõjul molekuli elektronpilve konfiguratsioon ja see sunnib seda teatud viisil reageerima. Sel viisil katalüüsib ensüüm reaktsiooni. Antikeha molekulidel on ka õõnsused, milles seonduvad mitmesugused võõrained ja muutuvad seeläbi kahjutuks. Mudel “lukk ja võti”, mis selgitab valkude vastasmõju teiste ühenditega, võimaldab mõista ensüümide ja antikehade spetsiifilisust, s.o. nende võime reageerida ainult teatud ühenditega.
Valgud erinevat tüüpi organismides. Sarnase konfiguratsiooniga on ka valgud, mis täidavad erinevatel taime- ja loomaliikidel sama funktsiooni ning kannavad seetõttu sama nime. Kuid need erinevad mõnevõrra oma aminohappejärjestuse poolest. Kuna liigid lahknevad ühisest esivanemast, asendatakse teatud positsioonides mõned aminohapped teiste mutatsioonidega. Pärilikke haigusi põhjustavad kahjulikud mutatsioonid elimineeritakse loodusliku valiku teel, kuid kasulikud või vähemalt neutraalsed mutatsioonid võivad püsida. Mida lähemal on kaks liiki teineteisele, seda vähem on nende valkudes erinevusi.Mõned valgud muutuvad suhteliselt kiiresti, teised on väga konserveerunud. Viimaste hulka kuulub näiteks tsütokroom Koos hingamisteede ensüüm, mida leidub enamikus elusorganismides. Inimestel ja šimpansitel on selle aminohappejärjestus identne ja tsütokroomis Koos Nisus olid ainult 38% aminohapetest erinevad. Isegi kui võrrelda inimesi ja baktereid, siis tsütokroomide sarnasus Koos(erinevused mõjutavad siin 65% aminohapetest) on siiani näha, kuigi bakterite ja inimeste ühine esivanem elas Maal umbes kaks miljardit aastat tagasi. Tänapäeval kasutatakse fülogeneetilise (sugu-)puu koostamiseks sageli aminohappejärjestuste võrdlemist, mis peegeldab erinevate organismide vahelisi evolutsioonilisi suhteid.
Denatureerimine. Sünteesitud valgu molekul, voltimine, omandab oma iseloomuliku konfiguratsiooni. Seda konfiguratsiooni saab aga hävitada kuumutamisel, pH muutmisel, kokkupuutel orgaaniliste lahustitega ja isegi lahuse lihtsalt loksutades, kuni selle pinnale ilmuvad mullid. Sel viisil modifitseeritud valku nimetatakse denatureeritud; see kaotab oma bioloogilise aktiivsuse ja muutub tavaliselt lahustumatuks. Tuntud näited denatureeritud valkudest on keedetud munad või vahukoor. Väikesed valgud, mis sisaldavad vaid umbes sadat aminohapet, on võimelised renatureerima, s.t. hankige uuesti algne konfiguratsioon. Kuid enamik valke muutub lihtsalt sassis polüpeptiidahelate massiks ega taasta oma eelmist konfiguratsiooni.Üks peamisi raskusi aktiivsete valkude eraldamisel on nende äärmine tundlikkus denaturatsiooni suhtes. See valkude omadus leiab kasulikku rakendust toiduainete säilitamisel: kõrge temperatuur denatureerib pöördumatult mikroorganismide ensüüme ja mikroorganismid surevad.
VALGU SÜNTEES Valkude sünteesimiseks peab elusorganismis olema ensüümide süsteem, mis on võimeline ühendama ühe aminohappe teisega. Teabeallikat on vaja ka selleks, et määrata, milliseid aminohappeid tuleks kombineerida. Kuna kehas on tuhandeid valke ja igaüks neist koosneb keskmiselt mitmesajast aminohappest, peab vajalik teave olema tõeliselt tohutu. See on salvestatud (sarnaselt sellele, kuidas salvestist salvestatakse magnetlindile) nukleiinhappemolekulides, mis moodustavad geenid. cm . ka PÄRILIKUD; NUKLEIINHAPPED.Ensüümide aktiveerimine. Aminohapetest sünteesitud polüpeptiidahel ei ole alati lõplikul kujul valk. Paljud ensüümid sünteesitakse esmalt mitteaktiivsete lähteainetena ja muutuvad aktiivseks alles pärast seda, kui teine ensüüm eemaldab ahela ühest otsast mitu aminohapet. Mõned seedeensüümid, näiteks trüpsiin, sünteesitakse sellel inaktiivsel kujul; need ensüümid aktiveeruvad seedetraktis ahela terminaalse fragmendi eemaldamise tulemusena. Hormooninsuliin, mille molekul aktiivsel kujul koosneb kahest lühikesest ahelast, sünteesitakse ühe ahela kujul, nn. proinsuliin. Seejärel eemaldatakse selle ahela keskmine osa ja ülejäänud fragmendid seostuvad omavahel, moodustades aktiivse hormooni molekuli. Komplekssed valgud tekivad alles pärast seda, kui valgu külge on kinnitunud konkreetne keemiline rühm ja see kinnitus nõuab sageli ka ensüümi.Metaboolne vereringe. Pärast looma söötmist süsiniku, lämmastiku või vesiniku radioaktiivsete isotoopidega märgistatud aminohapetega liidetakse märgis kiiresti tema valkudesse. Kui märgistatud aminohapped lakkavad kehasse sisenemast, hakkab märgise hulk valkudes vähenema. Need katsed näitavad, et saadud valgud ei säili organismis eluea lõpuni. Kõik need, välja arvatud üksikud erandid, on dünaamilises olekus, lagunedes pidevalt aminohapeteks ja seejärel uuesti sünteesides.Mõned valgud lagunevad, kui rakud surevad ja hävivad. See juhtub kogu aeg, näiteks soolestiku sisepinda vooderdavate punaste vereliblede ja epiteelirakkudega. Lisaks toimub elusrakkudes ka valkude lagunemine ja taassüntees. Kummalisel kombel teatakse valkude lagunemisest vähem kui nende sünteesist. Siiski on selge, et lagunemine hõlmab proteolüütilisi ensüüme, mis on sarnased ensüümidega, mis lagundavad valgud seedetraktis aminohapeteks.
Erinevate valkude poolväärtusaeg varieerub mitmest tunnist mitme kuuni. Ainus erand on kollageeni molekul. Kui need on moodustatud, jäävad need stabiilseks ja neid ei uuendata ega asendata. Aja jooksul aga muutuvad osad nende omadustest, eelkõige elastsus, ning kuna need ei uuene, siis selle tulemuseks on teatud vanusega seotud muutused, näiteks nahale tekivad kortsud.
Sünteetilised valgud. Keemikud on juba ammu õppinud aminohappeid polümeriseerima, kuid aminohappeid kombineeritakse ebakorrapäraselt, nii et sellise polümerisatsiooni saadused ei sarnane looduslike omadega. Tõsi, aminohappeid on võimalik teatud järjekorras kombineerida, mis võimaldab saada mõningaid bioloogiliselt aktiivseid valke, eriti insuliini. Protsess on üsna keeruline ja nii on võimalik saada ainult neid valke, mille molekulid sisaldavad sadakond aminohapet. Selle asemel on eelistatav sünteesida või isoleerida soovitud aminohappejärjestusele vastava geeni nukleotiidjärjestus ja seejärel sisestada see geen bakterisse, mis toodab replikatsiooni teel suures koguses soovitud produkti. Sellel meetodil on aga ka omad miinused. cm . ka GENEETIKA. VALK JA TOITUMINE Kui organismis olevad valgud lagundatakse aminohapeteks, saab neid aminohappeid uuesti kasutada valkude sünteesimiseks. Samal ajal lagunevad aminohapped ise, mistõttu neid täielikult uuesti ei kasutata. Samuti on selge, et kasvu, raseduse ja haavade paranemise ajal peab valkude süntees ületama lagunemise. Keha kaotab pidevalt mõningaid valke; Need on juuste, küünte ja naha pinnakihi valgud. Seetõttu peab iga organism valkude sünteesimiseks saama aminohappeid toidust. Rohelised taimed sünteesivad CO-st 2 , vesi ja ammoniaak või nitraadid on kõik 20 aminohapet, mida leidub valkudes. Paljud bakterid on samuti võimelised sünteesima aminohappeid suhkru (või mõne samaväärse) ja fikseeritud lämmastiku juuresolekul, kuid lõpuks tarnivad suhkrut rohelised taimed. Loomadel on piiratud võime sünteesida aminohappeid; nad saavad aminohappeid rohelisi taimi või muid loomi süües. Seedetraktis lagundatakse imendunud valgud aminohapeteks, viimased imenduvad ning neist ehitatakse üles antud organismile omased valgud. Ükski imendunud valk ei ole sellisena kehastruktuuridesse lülitatud. Ainus erand on see, et paljudel imetajatel võivad osad ema antikehad pääseda tervena läbi platsenta loote vereringesse ja emapiima kaudu (eriti mäletsejalistel) kanduda vastsündinule kohe pärast sündi.Valgu vajadus. On selge, et elu säilitamiseks peab organism toidust saama teatud koguse valku. Selle vajaduse ulatus sõltub aga mitmest tegurist. Keha vajab toitu nii energiaallikana (kalorid) kui ka materjalina oma struktuuride ülesehitamiseks. Energiavajadus on esikohal. See tähendab, et kui toidus on vähe süsivesikuid ja rasvu, kasutatakse toiduvalke mitte nende enda valkude sünteesiks, vaid kaloriallikana. Pikaajalise paastumise ajal kasutatakse energiavajaduse rahuldamiseks isegi teie enda valke. Kui toidus on piisavalt süsivesikuid, saab valgu tarbimist vähendada.Lämmastiku tasakaal. Keskmiselt u. 16% valkude kogumassist on lämmastik. Valkudes sisalduvate aminohapete lagundamisel eritub neis sisalduv lämmastik organismist uriiniga ja (vähemal määral) väljaheitega erinevate lämmastikuühendite kujul. Seetõttu on valgulise toitumise kvaliteedi hindamiseks mugav kasutada sellist indikaatorit nagu lämmastiku tasakaal, s.t. kehasse siseneva lämmastiku koguse ja ööpäevas erituva lämmastiku koguse vahe (grammides). Täiskasvanu normaalse toitumise korral on need kogused võrdsed. Kasvavas organismis on väljutatava lämmastiku hulk väiksem kui vastuvõetav, s.t. saldo on positiivne. Kui toidus on valgupuudus, on saldo negatiivne. Kui toidus on küllaldaselt kaloreid, aga valke selles pole, säästab organism valke. Samal ajal valkude ainevahetus aeglustub ning aminohapete korduv kasutamine valgusünteesis toimub võimalikult suure efektiivsusega. Kaod on aga vältimatud ning lämmastikuühendid erituvad endiselt uriiniga ja osaliselt ka väljaheitega. Valgupaastu ajal kehast päevas eritunud lämmastiku kogus võib olla igapäevase valgupuuduse mõõt. Loomulik on eeldada, et selle defitsiidiga võrdväärse valgukoguse lisamisega dieeti saab taastada lämmastiku tasakaalu. Siiski ei ole. Pärast sellise valgukoguse saamist hakkab organism aminohappeid vähem tõhusalt kasutama, mistõttu on lämmastiku tasakaalu taastamiseks vaja täiendavat valku.Kui valgu kogus toidus ületab lämmastiku tasakaalu säilitamiseks vajaliku, ei paista kahju olevat. Üleliigseid aminohappeid kasutatakse lihtsalt energiaallikana. Eriti ilmeka näitena tarbivad eskimod vähe süsivesikuid ja umbes kümme korda rohkem valku, mis on vajalik lämmastiku tasakaalu säilitamiseks. Enamasti ei ole valgu kasutamine energiaallikana siiski kasulik, sest teatud kogus süsivesikuid võib toota palju rohkem kaloreid kui sama kogus valku. Vaestes riikides saavad inimesed oma kalorid süsivesikutest ja tarbivad minimaalses koguses valku.
Kui organism saab vajaliku hulga kaloreid mittevalgutoodetena, siis minimaalne valgukogus lämmastiku tasakaalu säilimise tagamiseks on ca. 30 g päevas. Umbes nii palju valku sisaldab neli viilu saia või 0,5 liitrit piima. Tavaliselt peetakse optimaalseks veidi suuremat arvu; soovitatav 50-70 g.
Asendamatud aminohapped. Seni käsitleti valku tervikuna. Samal ajal peavad valkude sünteesi toimumiseks kehas olema kõik vajalikud aminohapped. Looma keha ise on võimeline sünteesima mõningaid aminohappeid. Neid nimetatakse asendatavateks, kuna need ei pea tingimata toidus olema, oluline on vaid, et üldine valguvaru lämmastikuallikana oleks piisav; siis, kui asendamatutest aminohapetest on puudus, saab organism neid sünteesida nende arvelt, mida leidub liigselt. Ülejäänud, "asendamatuid" aminohappeid ei saa sünteesida ja need tuleb organismi varustada toiduga. Inimestele hädavajalikud on valiin, leutsiin, isoleutsiin, treoniin, metioniin, fenüülalaniin, trüptofaan, histidiin, lüsiin ja arginiin. (Kuigi arginiini saab organismis sünteesida, klassifitseeritakse see asendamatuks aminohappeks, kuna seda ei toodeta piisavas koguses vastsündinutel ja kasvavatel lastel. Teisest küljest võivad mõned neist toidust saadavatest aminohapetest muutuda täiskasvanu jaoks ebavajalikuks inimene.)See asendamatute aminohapete loend on teistel selgroogsetel ja isegi putukatel ligikaudu sama. Valkude toiteväärtus määratakse tavaliselt kasvavatele rottidele söötmise ja loomade kaalutõusu jälgimise teel.
Valkude toiteväärtus. Valgu toiteväärtuse määrab asendamatu aminohape, millest on kõige rohkem puudus. Illustreerime seda näitega. Meie kehas olevad valgud sisaldavad keskmiselt u. 2% trüptofaani (massi järgi). Oletame, et dieet sisaldab 10 g valku, mis sisaldab 1% trüptofaani ja selles on piisavalt muid asendamatuid aminohappeid. Meie puhul võrdub 10 g seda mittetäielikku valku sisuliselt 5 g täisvalguga; ülejäänud 5 g saab olla ainult energiaallikas. Arvestage, et kuna aminohappeid organismis praktiliselt ei säilitata ja valkude sünteesi toimumiseks peavad kõik aminohapped olema samaaegselt olemas, saab asendamatute aminohapete tarbimise mõju tuvastada ainult siis, kui need kõik siseneda kehasse samal ajal. Enamiku loomsete valkude keskmine koostis on lähedane inimorganismi valkude keskmisele koostisele, seega on vähetõenäoline, et meil tekib aminohapete vaegus, kui meie toidus on rikkalikult toiduaineid, nagu liha, munad, piim ja juust. Siiski on valke, nagu želatiin (kollageeni denaturatsiooni produkt), mis sisaldavad väga vähe asendamatuid aminohappeid. Taimsed valgud, kuigi nad on selles mõttes paremad kui želatiin, on ka asendamatute aminohapete poolest vaesed; Nendes on eriti vähe lüsiini ja trüptofaani. Sellegipoolest ei saa puhtalt taimetoitlust üldse kahjulikuks pidada, välja arvatud juhul, kui see tarbib veidi suuremas koguses taimseid valke, mis on piisav, et varustada organismi asendamatute aminohapetega. Taimed sisaldavad kõige rohkem valku oma seemnetes, eriti nisu ja erinevate kaunviljade seemnetes. Valgurikkad on ka noored võrsed, näiteks spargel.Sünteetilised valgud toidus. Lisades mittetäielikele valkudele, näiteks maisivalkudele, väikeses koguses sünteetilisi asendamatuid aminohappeid või aminohapperikkaid valke, saab viimaste toiteväärtust oluliselt tõsta, s.t. suurendades seeläbi tarbitava valgu hulka. Teine võimalus on kasvatada baktereid või pärmi nafta süsivesinikel, lisades lämmastikuallikana nitraate või ammoniaaki. Sel viisil saadud mikroobne valk võib olla kodulindude või kariloomade söödaks või inimestele otse tarbitav. Kolmas, laialdaselt kasutatav meetod kasutab mäletsejaliste füsioloogiat. Mäletsejalistel mao algosas nn. Vatsas elavad bakterite ja algloomade erivormid, mis muudavad mittetäielikud taimsed valgud terviklikumateks mikroobseteks valkudeks, mis omakorda pärast seedimist ja imendumist loomseteks valkudeks. Karbamiidi, odavat sünteetilist lämmastikku sisaldavat ühendit, võib lisada loomasöödale. Vatsas elavad mikroorganismid kasutavad uurea lämmastikku, et muuta süsivesikud (mida on söödas palju rohkem) valkudeks. Umbes kolmandik kogu loomasöödas leiduvast lämmastikust võib tulla karbamiidi kujul, mis sisuliselt tähendab teatud määral valgu keemilist sünteesi. USA-s on sellel meetodil oluline roll ühe valgu hankimise viisina.KIRJANDUS Murray R., Grenner D., Mayes P., Rodwell W. Inimese biokeemia, vol. 12. M., 1993Alberts B, Bray D, Lewis J jt. Molekulaarrakubioloogia, vol. 13. M., 1994
Pilet 2. 1. Lipiidide olemuse olulised toitumistegurid. Mõned lipiidid ei sünteesita inimkehas ja on seetõttu olulised toitumistegurid. Nende hulka kuuluvad kahe või enama kaksiksidemega rasvhapped (polüeen) - asendamatud rasvhapped. Mõned neist hapetest on kohalike hormoonide – eikosanoidide – sünteesi substraadid (teema 8.10).
Rasvlahustuvad vitamiinid täidab erinevaid funktsioone: vitamiin A osaleb nii nägemise kui ka rakkude kasvu ja diferentseerumise protsessis; selle võime pärssida teatud tüüpi kasvajate kasvu on tõestatud; vitamiin K osaleb vere hüübimises; D-vitamiini osaleb kaltsiumi metabolismi reguleerimises; vitamiin E- antioksüdant, pärsib vabade radikaalide teket ja seega neutraliseerib lipiidide peroksüdatsioonist tingitud rakukahjustusi.
Dokument
2.Valkude struktuur ja struktuurse organiseerituse tasemed
Valkude struktuursel organiseerimisel on neli taset: primaarne, sekundaarne, tertsiaarne ja kvaternaarne. Igal tasemel on oma omadused.
Valgu esmane struktuur
Valkude põhistruktuur on aminohapete lineaarne polüpeptiidahel, mis on ühendatud peptiidsidemetega. Primaarstruktuur on valgu molekuli struktuurse organiseerituse kõige lihtsam tase. Kõrge stabiilsuse annavad sellele kovalentsed peptiidsidemed ühe aminohappe α-aminorühma ja teise aminohappe α-karboksüülrühma vahel. [saade].
Kui proliini või hüdroksüproliini iminorühm osaleb peptiidsideme moodustumisel, on sellel erinev vorm [saade].
Kui rakkudes tekivad peptiidsidemed, aktiveeritakse esmalt ühe aminohappe karboksüülrühm ja seejärel ühineb see teise aminohappe aminorühmaga. Polüpeptiidide laboratoorne süntees viiakse läbi ligikaudu samal viisil.
Peptiidside on polüpeptiidahela korduv fragment. Sellel on mitmeid omadusi, mis mõjutavad mitte ainult primaarstruktuuri kuju, vaid ka polüpeptiidahela kõrgemat organiseerituse taset:

koplanaarsus – kõik peptiidrühma kuuluvad aatomid on samal tasapinnal;
võime eksisteerida kahes resonantsvormis (keto- või enoolvorm);
asendajate trans-positsioon C-N sideme suhtes;
võime moodustada vesiniksidemeid ja iga peptiidrühm võib moodustada kaks vesiniksidet teiste rühmadega, sealhulgas peptiidrühmadega.
Erandiks on peptiidrühmad, mis hõlmavad proliini või hüdroksüproliini aminorühma. Nad on võimelised moodustama ainult ühe vesiniksideme (vt eespool). See mõjutab valgu sekundaarse struktuuri moodustumist. Polüpeptiidahel piirkonnas, kus proliin või hüdroksüproliin asub, paindub kergesti, kuna seda ei hoia, nagu tavaliselt, teine vesinikside.
Peptiidide ja polüpeptiidide nomenklatuur. Peptiidide nimi koosneb nende koostises olevate aminohapete nimedest. Kaks aminohapet moodustavad dipeptiidi, kolm moodustavad tripeptiidi, neli moodustavad tetrapeptiidi jne. Igal mis tahes pikkusega peptiidil või polüpeptiidahelal on N-terminaalne aminohape, mis sisaldab vaba aminorühma ja C-otsa aminohape, mis sisaldab vaba karboksüülrühma Grupp. Polüpeptiidide nimetamisel loetletakse kõik aminohapped järjestikku, alustades N-terminaalsest, asendades nende nimedes, välja arvatud C-otsa, järelliide -in -üüliga (kuna peptiidides sisalduvatel aminohapetel ei ole enam karboksüülrühm, vaid karbonüülrühm). Näiteks joonisel fig. 1 tripeptiid - leuc muda fenüülalaan muda treoon sisse.
Valgu esmase struktuuri tunnused. Polüpeptiidahela selgroos vahelduvad jäigad struktuurid (lamedad peptiidirühmad) suhteliselt liikuvate piirkondadega (-CHR), mis on võimelised sidemete ümber pöörlema. Sellised polüpeptiidahela struktuurilised omadused mõjutavad selle ruumilist paigutust.
Valkude sekundaarne struktuur
Sekundaarne struktuur on viis polüpeptiidahela voltimiseks järjestatud struktuuriks sama ahela peptiidrühmade või külgnevate polüpeptiidahelate vahel vesiniksidemete moodustumise tõttu. Konfiguratsiooni järgi jagunevad sekundaarsed struktuurid spiraalseteks (α-heeliks) ja kihilisteks volditud (β-struktuur ja rist-β-vorm).


α-heeliks. See on teatud tüüpi sekundaarne valgu struktuur, mis näeb välja nagu tavaline spiraal, mis on moodustunud peptiidsete vesiniksidemete tõttu ühes polüpeptiidahelas. α-heeliksi struktuuri mudeli (joonis 2), mis võtab arvesse kõiki peptiidsideme omadusi, pakkusid välja Pauling ja Corey. α-heeliksi peamised omadused:
spiraalse sümmeetriaga polüpeptiidahela spiraalne konfiguratsioon;
vesiniksidemete moodustumine iga esimese ja neljanda aminohappejäägi peptiidrühmade vahel;
spiraalsete pöörete korrapärasus;
kõigi α-heeliksi aminohappejääkide ekvivalentsus, sõltumata nende kõrvalradikaalide struktuurist;
aminohapete kõrvalradikaalid ei osale α-heeliksi moodustamises.
Väliselt näeb α-heeliks välja nagu elektripliidi kergelt venitatud spiraal. Esimese ja neljanda peptiidrühma vaheliste vesiniksidemete regulaarsus määrab polüpeptiidahela pöörete regulaarsuse. Ühe pöörde kõrgus ehk α-heeliksi samm on 0,54 nm; see sisaldab 3,6 aminohappejääki, st iga aminohappejääk liigub piki telge (ühe aminohappejäägi kõrgus) 0,15 nm (0,54:3,6 = 0,15 nm), mis võimaldab rääkida kõigi aminohappejääkide samaväärsusest α-heeliksis. α-heeliksi regulaarsusperiood on 5 pööret või 18 aminohappejääki; ühe perioodi pikkus on 2,7 nm. Riis. 3. Pauling-Corey a-helix mudel
β-struktuur. See on teatud tüüpi sekundaarne struktuur, millel on polüpeptiidahela veidi kõver konfiguratsioon ja mis on moodustatud peptiidsete vesiniksidemetega ühe polüpeptiidahela üksikutes osades või külgnevates polüpeptiidahelates. Seda nimetatakse ka kihiliseks voldikuks. β-struktuure on erinevaid. Valgu ühest polüpeptiidahelast moodustatud piiratud kihilisi piirkondi nimetatakse rist-β vormiks (lühike β struktuur). Polüpeptiidahela silmuste peptiidrühmade vahel moodustuvad rist-β vormis vesiniksidemed. Teine tüüp - täielik β-struktuur - on iseloomulik kogu polüpeptiidahelale, millel on piklik kuju ja mida hoiavad peptiidsidemed külgnevate paralleelsete polüpeptiidahelate vahel (joonis 3). See struktuur meenutab akordioni lõõtsa. Pealegi on võimalikud β-struktuuride variandid: need võivad olla moodustatud paralleelsete ahelatena (polüpeptiidahelate N-otsad on suunatud samas suunas) ja antiparalleelsetena (N-terminaalsed otsad on suunatud eri suundades). Ühe kihi külgradikaalid asetatakse teise kihi külgradikaalide vahele.
Valkudes on vesiniksidemete ümberkorraldamise tõttu võimalikud üleminekud α-struktuuridelt β-struktuuridele ja tagasi. Regulaarsete peptiidsete vesiniksidemete asemel piki ahelat (tänu millele polüpeptiidahel on keerdunud spiraaliks) kerivad spiraalsed lõigud lahti ja vesiniksidemed sulguvad polüpeptiidahelate piklike fragmentide vahel. Seda üleminekut leidub keratiinis, juuste valguses. Leeliseliste pesuvahenditega juukseid pestes hävib kergesti β-keratiini spiraalne struktuur ja see muutub α-keratiiniks (lokkis juuksed sirguvad).
Valkude regulaarsete sekundaarstruktuuride (α-heeliksid ja β-struktuurid) hävitamist nimetatakse analoogselt kristalli sulamisega polüpeptiidide “sulamiseks”. Sel juhul katkevad vesiniksidemed ja polüpeptiidahelad muutuvad juhusliku sasipuntra kujul. Järelikult määravad sekundaarstruktuuride stabiilsuse peptiidsete vesiniksidemed. Muud tüüpi sidemed selles peaaegu ei osale, välja arvatud disulfiidsidemed piki polüpeptiidahelat tsüsteiinijääkide asukohtades. Lühikesed peptiidid suletakse disulfiidsidemete tõttu tsükliteks. Paljud valgud sisaldavad nii α-spiraalseid piirkondi kui ka β-struktuure. 100% α-heeliksist koosnevaid looduslikke valke peaaegu pole (erandiks on paramüosiin, lihasvalk, mis koosneb 96-100% α-heeliksist), samas kui sünteetilistel polüpeptiididel on 100% heeliksit.
Teistel valkudel on erinev keerdumisaste. Paramüosiini, müoglobiini ja hemoglobiini puhul täheldatakse α-spiraalsete struktuuride suurt sagedust. Seevastu trüpsiinis, ribonukleaasis, on märkimisväärne osa polüpeptiidahelast volditud kihilisteks β-struktuurideks. Tugikudede valgud: keratiin (karva-, villavalk), kollageen (kõõluste, naha valk), fibroiin (loodusliku siidi valk) on polüpeptiidahelate β-konfiguratsiooniga. Valkude polüpeptiidahelate erinevad helilisuse astmed näitavad, et ilmselgelt on olemas jõud, mis osaliselt häirivad spiraali või "murvad" polüpeptiidahela korrapärase voltimise. Selle põhjuseks on valgu polüpeptiidahela kompaktsem voltimine teatud mahus, st tertsiaarseks struktuuriks.
Valkude tertsiaarne struktuur
Valgu tertsiaarne struktuur on viis, kuidas polüpeptiid ahel on ruumis paigutatud. Tertsiaarse struktuuri kuju järgi jagunevad valgud peamiselt kera- ja fibrillaarseteks. Kerakujulised valgud on enamasti ellipsoidse kujuga ja fibrillaarsed (niiditaolised) valgud on pikliku kujuga (varda või spindli kuju).
Valkude tertsiaarse struktuuri konfiguratsioon ei anna aga veel põhjust arvata, et fibrillaarsetel valkudel on ainult β-struktuur ja globulaarsetel valkudel on α-spiraalne struktuur. On fibrillaarseid valke, millel on spiraalne, mitte kihiline volditud sekundaarne struktuur. Näiteks α-keratiin ja paramüosiin (molluskite sulgurlihase valk), tropomüosiinid (skeletilihaste valgud) kuuluvad fibrillaarsete valkude hulka (on pulgakujuline) ja nende sekundaarne struktuur on α-heeliks; seevastu globulaarsed valgud võivad sisaldada suurt hulka β-struktuure.
Lineaarse polüpeptiidahela spiraliseerumine vähendab selle suurust ligikaudu 4 korda; ja tertsiaarsesse struktuuri pakkimine muudab selle kümneid kordi kompaktsemaks kui algne kett.
Sidemed, mis stabiliseerivad valgu tertsiaarset struktuuri. Aminohapete külgradikaalide vahelised sidemed mängivad rolli tertsiaarse struktuuri stabiliseerimisel. Need ühendused võib jagada järgmisteks osadeks:
tugev (kovalentne) [saade].
nõrk (polaarne ja van der Waals) [saade].
Arvukad sidemed aminohapete külgradikaalide vahel määravad valgu molekuli ruumilise konfiguratsiooni.
Valgu tertsiaarse struktuuri korralduse tunnused. Polüpeptiidahela tertsiaarse struktuuri konformatsiooni määravad selles sisalduvate aminohapete kõrvalradikaalide omadused (mis ei oma märgatavat mõju primaar- ja sekundaarstruktuuride tekkele) ning mikrokeskkond, st. keskkond. Voldituna kipub valgu polüpeptiidahel võtma energeetiliselt soodsa vormi, mida iseloomustab minimaalne vaba energia sisaldus. Seetõttu moodustavad mittepolaarsed R-rühmad, "vältides" vett, justkui valgu tertsiaarse struktuuri sisemise osa, kus asub põhiosa polüpeptiidahela hüdrofoobsetest jääkidest. Valgukerakese keskel pole veemolekule peaaegu üldse. Aminohappe polaarsed (hüdrofiilsed) R-rühmad asuvad selle hüdrofoobse tuuma välisküljel ja on ümbritsetud veemolekulidega. Polüpeptiidahel on kolmemõõtmelises ruumis keerukalt painutatud. Kui see paindub, on sekundaarne spiraalne konformatsioon häiritud. Ahel "katkeneb" nõrkades kohtades, kus asub proliin või hüdroksüproliin, kuna need aminohapped on ahelas liikuvamad, moodustades teiste peptiidrühmadega ainult ühe vesiniksideme. Teine paindekoht on glütsiin, millel on väike R-rühm (vesinik). Seetõttu kipuvad teiste aminohapete R-rühmad virnastatud kujul hõivama glütsiini asukohas vaba ruumi. Mitmed aminohapped – alaniin, leutsiin, glutamaat, histidiin – aitavad kaasa stabiilsete spiraalsete struktuuride säilimisele valkudes ning nagu metioniin, valiin, isoleutsiin, asparagiinhape soodustavad β-struktuuride teket. Tertsiaarse konfiguratsiooniga valgumolekulis on piirkonnad α-heeliksite (spiraalsed), β-struktuuride (kihilised) ja juhusliku mähise kujul. Ainult valgu õige ruumiline voltimine muudab selle aktiivseks; selle rikkumine toob kaasa muutused valgu omadustes ja bioloogilise aktiivsuse kadumise.
Kvaternaarne valgu struktuur
Ühest polüpeptiidahelast koosnevatel valkudel on ainult tertsiaarne struktuur. Nende hulka kuuluvad müoglobiin – lihaskoe valk, mis osaleb hapniku sidumises, mitmed ensüümid (lüsosüüm, pepsiin, trüpsiin jne). Mõned valgud on aga ehitatud mitmest polüpeptiidahelast, millest igaühel on tertsiaarne struktuur. Selliste valkude puhul on kasutusele võetud kvaternaarse struktuuri mõiste, mis kujutab endast mitme tertsiaarse struktuuriga polüpeptiidahela organiseerimist üheks funktsionaalseks valgu molekuliks. Sellist kvaternaarse struktuuriga valku nimetatakse oligomeeriks ja selle tertsiaarse struktuuriga polüpeptiidahelaid protomeerideks või subühikuteks (joonis 4).

Organisatsiooni kvaternaarsel tasemel säilitavad valgud tertsiaarse struktuuri põhikonfiguratsiooni (kerakujuline või fibrillaarne). Näiteks hemoglobiin on kvaternaarse struktuuriga valk, mis koosneb neljast alaühikust. Iga alaühik on globulaarne valk ja üldiselt on hemoglobiinil ka globulaarne konfiguratsioon. Juukse- ja villavalgud - keratiinid, mis on tertsiaarselt seotud fibrillaarsete valkudega, on fibrillaarse konformatsiooni ja kvaternaarse struktuuriga.
Valgu kvaternaarse struktuuri stabiliseerumine. Kõik valgud, millel on kvaternaarne struktuur, eraldatakse üksikute makromolekulide kujul, mis ei lagune subühikuteks. Allüksuste pindade vahelised kontaktid on võimalikud ainult aminohappejääkide polaarsete rühmade tõttu, kuna iga polüpeptiidahela tertsiaarse struktuuri moodustumisel saavad mittepolaarsete aminohapete külgradikaalid (mis moodustavad suurema osa ahelast). kõik proteinogeensed aminohapped) on peidetud subühiku sees. Nende polaarsete rühmade vahel moodustub arvukalt ioonseid (soola-), vesiniku- ja mõnel juhul disulfiidsidemeid, mis hoiavad alaühikuid kindlalt korrastatud kompleksina. Vesiniksidemeid purustavate või disulfiidsildu vähendavate ainete kasutamine põhjustab protomeeride lagunemist ja valgu kvaternaarse struktuuri hävimist. Tabelis 1 on kokku võetud andmed sidemete kohta, mis stabiliseerivad valgu molekuli erinevaid organiseerituse tasemeid [saade].
Mõnede fibrillaarsete valkude struktuurse korralduse tunnused
Fibrillaarsete valkude struktuursel ülesehitusel on võrreldes globulaarsete valkudega mitmeid tunnuseid. Neid omadusi võib näha keratiini, fibroiini ja kollageeni näitel. Keratiinid eksisteerivad α- ja β-konformatsioonis. α-keratiinidel ja fibroiinil on kihiline-volditud sekundaarne struktuur, kuid keratiinis on ahelad paralleelsed ja fibroiinis antiparalleelsed (vt joonis 3); Lisaks sisaldab keratiin ahelatevahelisi disulfiidsidemeid, samas kui fibroiinil neid pole. Disulfiidsidemete katkemine viib keratiinide polüpeptiidahelate eraldumiseni. Vastupidi, maksimaalse arvu disulfiidsidemete moodustumine keratiinides kokkupuutel oksüdeerivate ainetega loob tugeva ruumilise struktuuri. Üldiselt on fibrillaarsetes valkudes, erinevalt globulaarsetest valkudest, mõnikord raske eristada rangelt erinevaid organiseerituse tasemeid. Kui aktsepteerime (nagu globulaarse valgu puhul), et tertsiaarstruktuur moodustatakse ruumis ühe polüpeptiidahela paigutamisega ja kvaternaarstruktuur mitme ahelaga, siis fibrillaarsetes valkudes on juba sekundaarstruktuuri moodustumisel kaasatud mitu polüpeptiidahelat. . Fibrillaarse valgu tüüpiline näide on kollageen, mis on inimkehas üks enim leiduvaid valke (umbes 1/3 kõigi valkude massist). Seda leidub kudedes, millel on suur tugevus ja madal venitatavus (luud, kõõlused, nahk, hambad jne). Kollageenis on kolmandiku aminohappejääkidest glütsiin ja umbes veerand või veidi rohkem proliin või hüdroksüproliin.

Kollageeni eraldatud polüpeptiidahel (esmane struktuur) näeb välja nagu katkendlik joon. See sisaldab umbes 1000 aminohapet ja selle molekulmass on umbes 10 5 (joonis 5, a, b). Polüpeptiidahel koosneb korduvast aminohapete triost (triplet), mille koostis on järgmine: gly-A-B, kus A ja B on mis tahes muud aminohapped peale glütsiini (enamasti proliin ja hüdroksüproliin). Kollageeni polüpeptiidahelad (või α-ahelad) sekundaarsete ja tertsiaarsete struktuuride moodustumisel (joonis 5, c ja d) ei suuda toota tüüpilisi spiraalse sümmeetriaga α-heelikse. Proliin, hüdroksüproliin ja glütsiin (antiheelilised aminohapped) segavad seda. Seetõttu moodustavad kolm α-ahelat justkui keerdunud spiraale, nagu kolm niiti, mis keerduvad ümber silindri. Kolm spiraalset α-ahelat moodustavad korduva kollageenistruktuuri, mida nimetatakse tropokollageeniks (joonis 5d). Tropokollageen on oma organisatsioonis kollageeni tertsiaarne struktuur. Korrapäraselt piki ahelat vahelduvad proliini ja hüdroksüproliini lamedad rõngad annavad sellele jäikuse, nagu ka ahelatevahelised sidemed tropokollageeni α-ahelate vahel (sellepärast on kollageen venimisele vastupidav). Tropokollageen on sisuliselt kollageenfibrillide alaühik. Tropokollageeni subühikute paigutamine kollageeni kvaternaarsesse struktuuri toimub astmeliselt (joonis 5e).
Kollageenistruktuuride stabiliseerumine toimub tänu ahelatevahelistele vesinik-, ioon- ja van der Waalsi sidemetele ning vähesele hulgale kovalentsetele sidemetele.
Kollageeni α-ahelatel on erinev keemiline struktuur. On olemas erinevat tüüpi α 1 ahelaid (I, II, III, IV) ja α 2 ahelaid. Sõltuvalt sellest, millised α 1 - ja α 2 -ahelad osalevad tropokollageeni kolmeahelalise heeliksi moodustamises, eristatakse nelja tüüpi kollageeni:
esimene tüüp - kaks α 1 (I) ja üks α 2 ahelat;
teine tüüp - kolm α 1 (II) ahelat;
kolmas tüüp - kolm α 1 (III) ahelat;
neljas tüüp - kolm α 1 (IV) ahelat.
Kõige tavalisem kollageen on esimest tüüpi: seda leidub luukoes, nahas, kõõlustes; teist tüüpi kollageeni leidub kõhrekoes jne. Ühte tüüpi koes võib olla erinevat tüüpi kollageeni.
Kollageenistruktuuride korrastatud agregatsioon, nende jäikus ja inertsus tagavad kollageenikiudude kõrge tugevuse. Kollageenvalgud sisaldavad ka süsivesikute komponente, st need on valkude-süsivesikute kompleksid.
Kollageen on rakuväline valk, mida moodustavad kõigis elundites leiduvad sidekoerakud. Seetõttu tekivad kollageeni kahjustamise (või selle moodustumise katkemise) korral elundite sidekoe tugifunktsioonide mitmekordsed rikkumised.
Alfa-polüpeptiidahel lõpeb aminohapete kombinatsiooniga valiin-leutsiin ja beeta-polüpeptiid ahel lõpeb kombinatsiooniga valiin-histidiin-leutsiin. Alfa- ja beeta-polüpeptiidahelad hemoglobiini molekulis ei ole paigutatud lineaarselt, see on esmane struktuur. Molekulisiseste jõudude olemasolu tõttu on polüpeptiidahelad keerdunud valkudele tüüpilise alfa-heeliksi (sekundaarstruktuur) kujul. Alfa-heeliksi spiraal ise paindub ruumiliselt iga alfa- ja beeta-polüpeptiidahela jaoks, moodustades munaja kujuga põimikud (tertsiaarne struktuur). Polüpeptiidahelate alfa-heeliksi heeliksite üksikud osad on tähistatud ladina tähtedega A-st H-ni. Kõik neli tertsiaarset kumerat alfa- ja beeta-polüpeptiidahelat paiknevad ruumiliselt teatud suhtes – kvaternaarses struktuuris. Neid ühendavad mitte tõelised keemilised sidemed, vaid molekulidevahelised jõud.
Selgus, et inimestel on kolm peamist normaalse hemoglobiini tüüpi: embrüonaalne - U, loote - F ja täiskasvanu hemoglobiin - A. HbU (nimetatud sõna emakas algustähe järgi) esineb embrüos 7. ja 12. elunädala vahel, siis see kaob ja ilmub loote hemoglobiin, mis pärast kolmandat kuud on loote peamine hemoglobiin. Pärast seda ilmub ingliskeelse sõna "adult" algustähe järel järk-järgult tavaline täiskasvanu hemoglobiin, mida nimetatakse HbA-ks. Loote hemoglobiini hulk väheneb järk-järgult, nii et sünnihetkel on 80% hemoglobiinist HbA ja ainult 20% HbF. Pärast sündi loote hemoglobiinisisaldus langeb jätkuvalt ja 2–3 eluaastaks on see vaid 1–2%. Sama palju loote hemoglobiini on täiskasvanul. Täiskasvanu ja üle 3-aastaste laste puhul peetakse patoloogiliseks HbF-i kogust, mis ületab 2%.
Lisaks normaalsetele hemoglobiinitüüpidele on praegu teada üle 50 patoloogilise variandi. Neid nimetati esmakordselt ladina tähtedega. Täht B puudub hemoglobiinitüüpide tähistamisel, kuna see tähistas algselt HbS-i.
Hemoglobiin (Hb)- punastes verelibledes esinev kromoproteiin, mis osaleb hapniku transportimisel kudedesse. Täiskasvanute hemoglobiini nimetatakse hemoglobiiniks A (Hb A). Selle molekulmass on umbes 65 000 Da. Hb A molekulil on kvaternaarne struktuur ja see sisaldab nelja subühikut – polüpeptiidahelaid (tähistatud α1, α2, β1 ja β2, millest igaüks on seotud heemiga.
Pidage meeles, et hemoglobiin on allosteeriline valk, mille molekulid võivad pöörduvalt muutuda ühest konformatsioonist teise. See muudab valgu afiinsust ligandide suhtes. Konformatsiooni, millel on ligandi suhtes väikseim afiinsus, nimetatakse pingeks ehk T-konformatsiooniks. Ligandi suhtes suurima afiinsusega konformatsiooni nimetatakse lõdvestunud ehk R-konformatsiooniks.
Hemoglobiini molekuli R- ja T-konformatsioonid on dünaamilises tasakaalus:
Erinevad keskkonnategurid võivad seda tasakaalu ühes või teises suunas nihutada. Allosteerilised regulaatorid, mis mõjutavad Hb afiinsust O2 suhtes, on: 1) hapnik; 2) H+ kontsentratsioon (keskmine pH); 3) süsinikdioksiid (CO2); 4) 2,3-difosfoglütseraat (DPG). Hapnikumolekuli kinnitumine ühele hemoglobiini subühikutest soodustab pingelise konformatsiooni üleminekut lõdvestunud konformatsioonile ja suurendab sama hemoglobiini molekuli teiste subühikute afiinsust hapniku suhtes. Seda nähtust nimetatakse koostööefektiks. Hemoglobiini hapnikuga seondumise kompleksne olemus kajastub hemoglobiini O2 küllastuskõveras, millel on S-kuju (joonis 3.1).

Joonis 3.1. Müoglobiini (1) ja hemoglobiini (2) hapnikuküllastuse kõverad.
Molekulaarsed vormid hemoglobiin erinevad üksteisest polüpeptiidahelate struktuuri poolest. Füsioloogilistes tingimustes eksisteeriva hemoglobiini mitmekesisuse näide on loote hemoglobiin (HbF), mis esineb veres inimese embrüonaalses arengujärgus. Erinevalt HbA-st sisaldab selle molekul 2 α- ja 2 γ-ahelat (st β-ahelad on asendatud γ-ahelatega). Sellisel hemoglobiinil on suurem afiinsus hapniku suhtes. Just see võimaldab embrüol saada hapnikku ema verest läbi platsenta. Varsti pärast sündi asendatakse lapse veres HbF HbA-ga.
Ebanormaalse või patoloogilise hemoglobiini näide on juba mainitud (vt 2.4.) hemoglobiin S, mida leidub sirprakulise aneemiaga patsientidel. Nagu te juba teate, erineb see hemoglobiinist A, asendades glutamaadi β-ahelates valiiniga. See aminohappe asendus põhjustab HbS vees lahustuvuse vähenemist ja selle afiinsuse vähenemist O2 suhtes.
Orgaanilised ained. Elusorganismid sisaldavad lisaks anorgaanilistele ka mitmesuguseid orgaanilisi aineid. Elusolendite orgaaniline aine koosneb peamiselt neljast keemilisest elemendist, mida nimetatakse biogeenne: süsinik, vesinik, hapnik ja lämmastik. Valkudes lisatakse nendele elementidele väävlit ja nukleiinhapetes fosforit.
Orgaanilise aine mitmekesisuse määrab suuresti süsinik. Tänu oma ainulaadsetele omadustele moodustab see element elu keemilise aluse. See võib moodustada kovalentseid sidemeid paljude aatomite ja nende rühmadega, moodustades ahelaid ja rõngaid, mis moodustavad erineva koostise, struktuuri, pikkuse ja kujuga orgaaniliste molekulide skeleti. Need omakorda moodustavad keerukaid keemilisi ühendeid, mis erinevad oma struktuuri ja funktsioonide poolest. Orgaaniliste molekulide mitmekesisuse peamiseks põhjuseks ei ole mitte niivõrd erinevus neid moodustavates aatomites, vaid pigem nende erinev järjestus molekulis.
Biopolümeeride kontseptsioon. Elusorganismis on orgaanilised ained esindatud kas väikeste, suhteliselt väikese molekulmassiga molekulide või suurte makromolekulidega. Madala molekulmassiga ühendite hulka kuuluvad aminohapped, suhkrud, orgaanilised happed, alkoholid, vitamiinid jne.
Valgud, polüsahhariidid ja nukleiinhapped on enamasti suure molekulmassiga struktuurid. Sellepärast neid kutsutaksegi makromolekulid(kreeka keelest makro- suur). Seega on enamiku valkude molekulmass vahemikus 5000 kuni 1 000 000 kõrgmolekulaarseid orgaanilisi ühendeid - valke, nukleiinhappeid, polüsahhariide, mille molekulid koosnevad suurest hulgast sama või erineva keemilise struktuuriga korduvatest ühikutest. biopolümeerid(kreeka keelest bios- elu ja poliitika- arvukalt). Lihtsaid molekule, millest biopolümeerid koosnevad, nimetatakse monomeerid. Valkude monomeerid on aminohapped, polüsahhariidide monomeerid on monosahhariidid ja nukleiinhapete monomeerid on nukleotiidid. Makromolekulid moodustavad umbes 90% raku kuivmassist.
Selles peatükis vaadeldakse kõiki kolme makromolekulide klassi ja nende monomeerühikuid. Arvesse on lisatud lipiidid - molekulid, mis on reeglina biopolümeeridest palju väiksemad, kuid täidavad ka kehas funktsioone.
Orgaaniliste ainete erirühm on bioloogiliselt aktiivsed ained: ensüümid, hormoonid, vitamiinid jne Need on struktuurilt mitmekesised; mõjutavad ainevahetust ja energia muundamist.
Erinevate organismirühmade rakkudes on teatud orgaaniliste ühendite sisaldus erinev. Näiteks loomarakkudes on ülekaalus valgud ja rasvad, taimerakkudes aga süsivesikud. Kuid erinevates rakkudes täidavad teatud orgaanilised ühendid sarnaseid funktsioone.
Oravad. Elusorganismides, makromolekulide hulgas, mängivad valgud oma funktsionaalse tähtsuse osas juhtivat rolli. Valgud on paljudes organismides domineerivad ja kvantitatiivsed. Loomade kehas moodustavad nad seega 40–50%, taimede kehas 20–35% nende kuivmassist. Valgud on heteropolümeerid, mille monomeerideks on aminohapped.
Aminohapped on valgumolekulide "ehituskivid". Aminohapped - orgaanilised ühendid, mis sisaldavad nii aminorühma (–NH), mida iseloomustavad aluselised omadused, kui ka happeliste omadustega karboksüülrühma (–COOH). Aminorühm ja karboksüülrühmad on ühendatud sama süsinikuaatomiga (joonis). Selle tunnuse järgi on kõik aminohapped üksteisega sarnased. Enamikul valku moodustavatel aminohapetel on üks karboksüülrühm ja üks aminorühm; neid aminohappeid nimetatakse neutraalne.
Osa molekulist, mida nimetatakse radikaalne (R) erinevatel aminohapetel on erinev struktuur (joonis). Erinevate aminohapete radikaal võib olla mittepolaarne või polaarne (laetud või laenguta), hüdrofoobne või hüdrofiilne, mis annab valkudele teatud omadused. Lisaks neutraalsetele on olemas aluselised aminohapped- rohkem kui ühe aminorühmaga, samuti happelised aminohapped- rohkem kui ühe karboksüülrühmaga. Täiendava amino- või hüdroksüülrühma olemasolu mõjutab radikaali omadusi. Valgu ruumilise struktuuri kujunemisel mängivad määravat rolli kõik aminohapperadikaalide omadused.
Teadaolevate aminohapete koguarv on umbes 200 ja ainult 20 tüüpi on seotud looduslike valkude moodustumisega. Neid aminohappeid nimetatakse valku moodustav(Tabel 2; tabelis on näidatud aminohapete täis- ja lühendatud nimetused, mitte meeldejätmiseks).
Tabel 2. Aluselised aminohapped ja nende lühendid
Taimed ja bakterid suudavad fotosünteesi esmastest saadustest sünteesida kõiki neile vajalikke aminohappeid. Inimene ja loom ei ole võimeline kõiki aminohappeid sünteesima, mistõttu nn asendamatud aminohapped need tuleks kätte saada valmis kujul koos toiduga.
Inimese jaoks asendamatud aminohapped on: lüsiin, valiin, leutsiin, isoleutsiin, treoniin, fenüülalaniin, trüptofaan Ja metioniin; lastele on samuti asendamatud arginiin Ja histidiin. Toiduvalke, mis sisaldavad kõiki asendamatuid aminohappeid, nimetatakse täieõiguslik, Erinevalt puudega, millel puuduvad teatud asendamatud aminohapped.
Nii aluseliste kui happeliste rühmade olemasolu ühes aminohappes määrab nende amfoteersuse ja kõrge reaktsioonivõime. Aminorühm
Ühe aminohappe (–NH 2) on võimeline interakteeruma teise aminohappe karboksüülrühmaga (–COOH), vabastades veemolekuli. Saadud molekul on dipeptiid (joonis.) ja nimetatakse –CO-NH– sidet peptiid. Dipeptiidimolekuli ühes otsas on vaba aminorühm ja teises karboksüülrühm. Tänu sellele saab dipeptiid enda külge siduda teisi aminohappeid, moodustades oligopeptiidid. Kui sel viisil on ühendatud palju aminohappeid (üle kümne), moodustub pikk ahel - polüpeptiid.
Peptiidid mängivad kehas olulist rolli. Paljud oligo- ja polüpeptiidid on hormoonid, antibiootikumid ja toksiinid.
Oligopeptiidide hulka kuuluvad näiteks hüpofüüsi hormoonid oksütotsiin ja vasopressiin, aga ka bradükiniin (valupeptiid) ja mõned opiaadid (inimeste "looduslikud ravimid"), mis toimivad valuvaigistitena. Regulaarne kasutamine Narkootikumide tarvitamine on väga ohtlik, see hävitab organismi opiaatide süsteemi, mistõttu narkodoosita sõltlane kogeb tugevat valu – “võõrutust”. Oligopeptiidide hulka kuuluvad mõned antibiootikumid, näiteks gramitsidiin S.
Polüpeptiidideks on ka hormoonid (insuliin, adrenokortikotroopne hormoon jne), antibiootikumid (gramitsidiin A), toksiinid (difteeriatoksiin).
Polüpeptiidahelad võivad olla väga pikad ja sisaldada mitmesuguseid aminohapete kombinatsioone. Polüpeptiide, mille molekul sisaldab 50 kuni mitu tuhat aminohappejääki molekulmassiga üle 6000, nimetatakse valkudeks.
Iga konkreetset valku iseloomustab rangelt konstantne aminohappejääkide koostis ja järjestus.
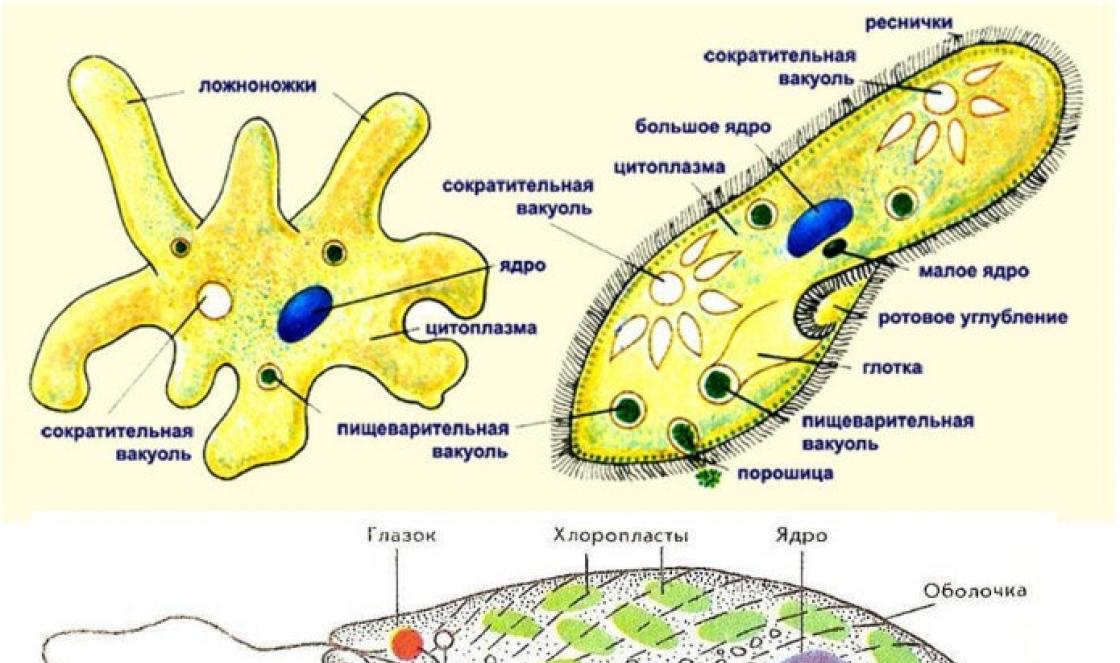
Valgu molekuli organiseerituse tasemed. Valgu molekulid võivad võtta erinevaid ruumilisi vorme – kehaehitus, mis esindavad nende organisatsiooni nelja taset (joon.)
Paljude peptiidsidemetega ühendatud aminohappejääkide ahel on esmane struktuur valgu molekul. See on kõige olulisem struktuur, kuna see määrab selle vormi, omadused ja funktsioonid. Põhistruktuurist lähtuvalt luuakse teist tüüpi struktuure. Just see struktuur on kodeeritud DNA molekulis. Igal üksikul kehavalgul on ainulaadne esmane struktuur. Kõigil konkreetse üksiku valgu (näiteks albumiini) molekulidel on sama aminohappejääkide vaheldumine, mis eristab albumiini mis tahes muust üksikust valgust. Primaarstruktuuri mitmekesisuse määrab polüpeptiidahela aminohappejääkide koostis, hulk ja järjestus.
Sekundaarne struktuur valgud tekivad vesiniksidemete moodustumisel polüpeptiidahela erinevate aminohappejääkide NH-rühmade vesinikuaatomi ja CO-rühmade hapnikuaatomi vahel. Polüpeptiidahel on keeratud spiraaliks. Kuigi vesiniksidemed on nõrgad, tagavad nad oma märkimisväärse arvu tõttu selle struktuuri stabiilsuse. Keratiini valgu molekulid on täiesti spiraalse konfiguratsiooniga. See on karvade, karusnaha, küüniste, sulgede ja sarvede struktuurne valk; see on osa selgroogsete naha väliskihist. Lisaks keratiinile on spiraalne sekundaarstruktuur iseloomulik fibrillaarsetele (niititaolistele) valkudele, nagu müosiin, fibrinogeen ja kollageen.
Valgu sekundaarset struktuuri saab lisaks heeliksile kujutada volditud kihina. Volditud kihis asetsevad paralleelselt mitu polüpeptiidahelat (või ühe polüpeptiidahela lõiku), mis moodustavad lameda konfiguratsiooni, mis on volditud nagu akordion (joonis b6). Näiteks valkfibroiinil, mis moodustab kookonite kudumise käigus siidiussi rööviku siidi sekreteerivate näärmete poolt eritatava põhiosa siidikiust, on sekundaarne struktuur volditud kihi kujul.
Tertsiaarne struktuur tekib S-S sidemete ("disulfiidsillade") vaheliste tsüsteiinijääkide (väävlit sisaldav aminohape), samuti vesiniku, ioonsete ja muude interaktsioonide kaudu. Tertsiaarne struktuur määrab valgu molekulide spetsiifilisuse ja nende bioloogilise aktiivsuse. Valkudel nagu müoglobiin (lihastes leiduv valk, mis osaleb hapnikuvarude loomises), trüpsiin (sooles leiduvaid valke lagundav ensüüm) on tertsiaarse struktuuriga.
Mõnel juhul ühendatakse mitu tertsiaarse struktuuriga polüpeptiidahelat üheks kompleksiks, moodustades seeläbi kvaternaarne struktuur. Selles ei ole valgu subühikud kovalentselt seotud ning tugevuse tagab nõrkade molekulidevaheliste jõudude koosmõju. Näiteks kvaternaarne struktuur on iseloomulik hemoglobiinivalgule, mis koosneb neljast valgu subühikust ja mittevalgulisest osast - heemist.
s 1. Mis on valgud? 2. Mis on valkude struktuur? 3. Mis on aminohapped? 4. Kuidas on aminohapped seotud polüpeptiidahela moodustamiseks? 5. Millised on valkude struktuurse organiseerituse tasemed? 6. Millised keemilised sidemed määravad valgu molekulide struktuurse organiseerituse erinevad tasemed? 7. A.B.C. on kolme tüüpi aminohappeid. Mitu viiest aminohappest koosneva polüpeptiidahela varianti saab konstrueerida? Kas polüpeptiididel on samad omadused?
Lühike kirjeldus:
Õpiku fragment: Bioloogiline keemia koos harjutuste ja ülesannetega: õpik / toim. Vastav liige RAMS S.E. Severina. M.: GEOTAR-Media, 2011. - 624 lk.: ill. 1. MOODUL: VALKUDE STRUKTUUR, OMADUSED JA FUNKTSIOONID
1. MOODUL: VALKUDE STRUKTUUR, OMADUSED JA FUNKTSIOONID
Mooduli struktuur | Teemad |
Moodulüksus 1 | 1.1. Valkude struktuurne korraldus. Loodusliku valgu konformatsiooni kujunemise etapid 1.2. Valkude funktsioneerimise alused. Ravimid kui ligandid, mis mõjutavad valgu funktsiooni 1.3. Valkude denatureerimine ja nende spontaanse renaturatsiooni võimalus |
Moodulüksus 2 | 1.4. Oligomeersete valkude struktuuri ja toimimise tunnused hemoglobiini näitel 1.5. Loodusliku valgu konformatsiooni säilitamine rakutingimustes 1.6. Valkude mitmekesisus. Valguperekonnad immunoglobuliinide näitel 1.7. Valkude füüsikalis-keemilised omadused ja nende eraldamise meetodid |
Moodulüksus 1 MONOMEERSETE VALKUDE STRUKTUURNE KORRALDUS JA NENDE TOIMIMISE ALUSED
Õppeeesmärgid Oskama:
1. Kasutada teadmisi valkude struktuurilistest iseärasustest ja valkude funktsioonide sõltuvusest nende struktuurist, et mõista pärilike ja omandatud proteinopaatiate tekkemehhanisme.
2. Selgitage mõnede ravimite terapeutilise toime mehhanisme kui ligandid, mis interakteeruvad valkudega ja muudavad nende aktiivsust.
3. Kasutage teadmisi valkude struktuuri ja konformatsioonilise labiilsuse kohta, et mõista nende struktuurset ja funktsionaalset ebastabiilsust ning kalduvust denatureerida muutuvates tingimustes.
4. Selgitage denatureerivate ainete kasutamist meditsiiniliste materjalide ja instrumentide steriliseerimiseks, samuti antiseptikumidena.
Tea:
1. Valkude struktuurse organiseerituse tasemed.
2. Valkude esmase struktuuri tähtsus, mis määrab nende struktuurse ja funktsionaalse mitmekesisuse.
3. Valkude aktiivtsentri moodustumise mehhanism ja selle spetsiifiline interaktsioon ligandiga, mis on valkude funktsioneerimise aluseks.
4. Näited eksogeensete ligandide (ravimid, toksiinid, mürgid) mõjust valkude konformatsioonile ja funktsionaalsele aktiivsusele.
5. Valkude denaturatsiooni põhjused ja tagajärjed, denaturatsiooni põhjustavad tegurid.
6. Näiteid denatureerivate faktorite kasutamisest meditsiinis antiseptikuna ja meditsiiniinstrumentide steriliseerimiseks.
TEEMA 1.1. VALGIDE STRUKTUURNE KORRALDUS. EMAKEELE TEKKE ETAPID
VALGUKONFORMATSIOONID
Oravad - need on heteropolümeerid molekulid (st koosneb mitmesugustest monomeeridest). Valgu monomeerid on 20 tüüpi α-aminohappeid, mis on omavahel ühendatud peptiidsidemetega.
Valgus sisalduvate aminohapete komplekti ja kombinatsiooni määrab vastavate geenide struktuur indiviidide DNA-s. Iga valk täidab vastavalt oma spetsiifilisele struktuurile oma funktsiooni. Antud organismi valkude kogum ( proteoom) määrab selle fenotüübilised omadused, samuti pärilike haiguste esinemise või eelsoodumuse nende tekkeks.
1. Aminohapped, millest koosnevad valgud. Peptiidside.
Valgud on heteropolümeerid, mis on ehitatud monomeeridest – 20 α-aminohappest.
Aminohapete üldvalem on esitatud allpool.

Aminohapped erinevad α-süsiniku aatomiga seotud radikaalide struktuuri, suuruse ja füüsikalis-keemiliste omaduste poolest. Aminohapete funktsionaalsed rühmad määravad erinevate α-aminohapete omaduste omadused. α-aminohapetes leiduvad radikaalid võib jagada mitmeks rühmaks:
proliin, Erinevalt ülejäänud 19 valgu monomeerist ei ole see aminohape, vaid proliinis olev radikaal on seotud nii α-süsiniku aatomiga kui ka iminorühmaga

Aminohapete lahustuvus vees on erinev. See on tingitud radikaalide võimest suhelda veega (hüdraat).
TO hüdrofiilsed Termin "laenguta" hõlmab radikaale, mis sisaldavad anioonseid, katioonseid ja polaarseid laenguta funktsionaalrühmi.
TO hüdrofoobne Termin "radikaalid" hõlmavad metüülrühmi, alifaatseid ahelaid või ringe sisaldavaid radikaale.
2. Peptiidsidemed ühendavad aminohappeid, moodustades peptiidid. Peptiidide sünteesi käigus interakteerub ühe aminohappe α-karboksüülrühm teise aminohappe α-aminorühmaga, moodustades peptiidside:
Valgud on polüpeptiidid, st. α-aminohapete lineaarsed polümeerid, mis on ühendatud peptiidsidemega (joonis 1.1.)
 Riis. 1.1. Peptiidide struktuuri kirjeldamiseks kasutatavad terminid
Riis. 1.1. Peptiidide struktuuri kirjeldamiseks kasutatavad terminid
Polüpeptiide moodustavate aminohapete monomeere nimetatakse aminohappejäägid. Korduvate rühmade ahel - NH-CH-CO- vormid peptiidi selgroog. Aminohappejääki, millel on vaba α-aminorühm, nimetatakse N-terminaalseks ja aminohappejääki, millel on vaba α-karboksüülrühm, C-terminaalseks. Peptiidid kirjutatakse ja loetakse N-otsast C-otsani.
Proliini iminorühma moodustatud peptiidside erineb teistest peptiidsidemetest: peptiidrühma lämmastikuaatomil puudub vesinik,
selle asemel tekib side radikaaliga, mille tulemusena on tsükli üks pool kaasatud peptiidi karkassi:

Peptiidid erinevad aminohapete koostise, aminohapete arvu ja aminohapete seose järjekorra poolest, näiteks Ser-Ala-Glu-Gis ja His-Glu-Ala-Ser on kaks erinevat peptiidi.
Peptiidsidemed on väga tugevad ja nende keemiline mitteensümaatiline hüdrolüüs nõuab karme tingimusi: analüüsitavat valku hüdrolüüsitakse kontsentreeritud vesinikkloriidhappes temperatuuril umbes 110° 24 tundi. Elus rakus võivad peptiidsidemed katkeda proteolüütilised ensüümid, helistas proteaasid või peptiidi hüdrolaasid.
3. Valkude esmane struktuur. Erinevate valkude peptiidahelates olevad aminohappejäägid ei vaheldu juhuslikult, vaid paiknevad kindlas järjekorras. Polüpeptiidahela aminohappejääkide lineaarset järjestust või vaheldumise järjekorda nimetatakse valgu esmane struktuur.
Iga üksiku valgu esmane struktuur on kodeeritud DNA molekulis (piirkonnas, mida nimetatakse geeniks) ja see realiseerub transkriptsiooni (info kopeerimine mRNA-le) ja translatsiooni (valgu esmase struktuuri süntees) käigus. Järelikult on üksiku inimese valkude esmane struktuur vanematelt lastele pärilikult edastatav informatsioon, mis määrab antud organismi valkude ehituslikud iseärasused, millest sõltub olemasolevate valkude talitlus (joon. 1.2.).
 Riis. 1.2. Genotüübi ja indiviidi kehas sünteesitud valkude konformatsiooni vaheline seos
Riis. 1.2. Genotüübi ja indiviidi kehas sünteesitud valkude konformatsiooni vaheline seos
Igal inimese kehas olevast ligikaudu 100 000 individuaalsest valgust on ainulaadne esmane struktuur. Sama tüüpi valgu (näiteks albumiini) molekulidel on sama aminohappejääkide vaheldumine, mis eristab albumiini mis tahes muust üksikust valgust.
Peptiidahela aminohappejääkide järjestust võib pidada teabe salvestamise vormiks. See teave määrab lineaarse peptiidahela ruumilise paigutuse kompaktsemaks kolmemõõtmeliseks struktuuriks, mida nimetatakse konformatsioon orav. Funktsionaalselt aktiivse valgu konformatsiooni moodustumise protsessi nimetatakse kokkuklapitavad
4. Valgu konformatsioon. Vaba pöörlemine peptiidi karkassis on võimalik peptiidrühma lämmastikuaatomi ja naabruses asuva α-süsiniku aatomi vahel, samuti α-süsiniku aatomi ja karbonüülrühma süsiniku vahel. Aminohappejääkide funktsionaalrühmade vastasmõju tõttu võib valkude primaarstruktuur omandada keerukamaid ruumilisi struktuure. Globulaarsetes valkudes on peptiidahelate konformatsiooni voltimisel kaks peamist taset: teisejärguline Ja tertsiaarne struktuur.
Valkude sekundaarne struktuur on ruumiline struktuur, mis on tekkinud peptiidi karkassi funktsionaalrühmade -C=O ja -NH- vahelise vesiniksidemete moodustumise tulemusena. Sel juhul võib peptiidahel omandada kahte tüüpi korrapäraseid struktuure: α-heeliksid Ja β-struktuurid.
IN α-heeliksid karbonüülrühma hapnikuaatomi ja 4. aminohappe amiidlämmastiku vesiniku vahel tekivad vesiniksidemed; aminohappejääkide külgahelad
paiknevad piki spiraali perifeeriat, osalemata sekundaarstruktuuri moodustamisel (joonis 1.3.).
Hulgiradikaalid ehk võrdset laengut kandvad radikaalid takistavad α-heeliksi moodustumist. Ringstruktuuriga proliinijääk katkestab α-heeliksi, kuna peptiidahela lämmastikuaatomi vesiniku puudumise tõttu ei saa vesiniksidemeid moodustada. Side lämmastiku ja α-süsiniku aatomi vahel on osa proliinitsüklist, mistõttu peptiidi karkass muutub selles punktis painutatud.
β-struktuur moodustub ühe polüpeptiidahela peptiidse karkassi lineaarsete piirkondade vahel, moodustades seeläbi volditud struktuure. Võib moodustuda polüpeptiidahelaid või nende osi paralleelselt või antiparalleelsed β-struktuurid. Esimesel juhul langevad interakteeruvate peptiidahelate N- ja C-otsad kokku, teisel juhul on need vastupidise suunaga (joon. 1.4).
Riis. 1.3. Valgu sekundaarne struktuur - α-heeliks
 Riis. 1.4. Paralleelsed ja antiparalleelsed β-lehtstruktuurid
Riis. 1.4. Paralleelsed ja antiparalleelsed β-lehtstruktuurid
β-struktuurid on tähistatud laiade nooltega: A - Antiparalleelne β-struktuur. B – Paralleelsed β-lehtstruktuurid
Mõnedes valkudes võivad β-struktuurid tekkida erinevate polüpeptiidahelate peptiidse karkassi aatomite vahel vesiniksidemete moodustumise tõttu.
Leidub ka valkudes ebaregulaarse sekundaarse piirkonnaga struktuur, mis hõlmab polüpeptiidi selgroo painutusi, silmuseid ja pöördeid. Need paiknevad sageli kohtades, kus peptiidahela suund muutub, näiteks paralleelse β-lehe struktuuri moodustumisel.
α-heeliksite ja β-struktuuride olemasolu põhjal võib globulaarsed valgud jagada nelja kategooriasse.
Riis. 1.5. Müoglobiini (A) ja hemoglobiini β-ahela (B) sekundaarne struktuur, mis sisaldab kaheksat α-heeliksi

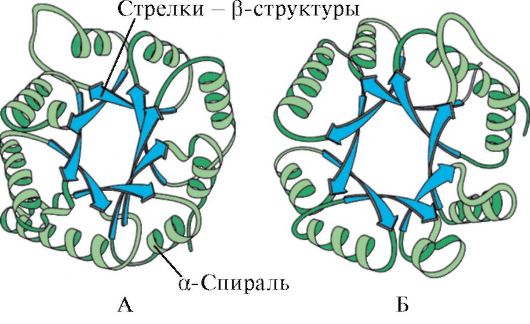 Riis. 1.6. Triosefosfaadi isomeraasi ja püruvaatkinaasi domeeni sekundaarne struktuur
Riis. 1.6. Triosefosfaadi isomeraasi ja püruvaatkinaasi domeeni sekundaarne struktuur
 Riis. 1.7. Immunoglobuliini (A) ja ensüümi superoksiiddismutaasi (B) konstantse domeeni sekundaarne struktuur
Riis. 1.7. Immunoglobuliini (A) ja ensüümi superoksiiddismutaasi (B) konstantse domeeni sekundaarne struktuur
IN neljas kategooria hõlmasid valke, mis sisaldavad väikeses koguses regulaarseid sekundaarstruktuure. Nende valkude hulka kuuluvad väikesed tsüsteiinirikkad valgud või metalloproteiinid.
Valkude tertsiaarne struktuur- teatud tüüpi konformatsioon, mis moodustub aminohapete radikaalide vastastikmõjude tõttu, mis võivad paikneda peptiidahelas üksteisest märkimisväärsel kaugusel. Enamik valke moodustab gloobulit meenutava ruumilise struktuuri (globulaarsed valgud).
Kuna hüdrofoobsed aminohapperadikaalid kipuvad ühinema läbi nn hüdrofoobsed interaktsioonid ja molekulidevaheliste van der Waalsi jõudude mõjul moodustub valgugloobuli sees tihe hüdrofoobne tuum. Hüdrofiilsed ioniseeritud ja ioniseerimata radikaalid paiknevad peamiselt valgu pinnal ja määravad selle lahustuvuse vees.
 Riis. 1.8. Sidemete tüübid, mis tekivad aminohappe radikaalide vahel valgu tertsiaarse struktuuri moodustumisel
Riis. 1.8. Sidemete tüübid, mis tekivad aminohappe radikaalide vahel valgu tertsiaarse struktuuri moodustumisel
1 - ioonne side- esineb positiivse ja negatiivse laenguga funktsionaalrühmade vahel;
2 - vesinikside- esineb hüdrofiilse laenguta rühma ja mis tahes muu hüdrofiilse rühma vahel;
3 - hüdrofoobsed interaktsioonid- tekivad hüdrofoobsete radikaalide vahel;
4 - disulfiidside- moodustub tsüsteiinijääkide SH-rühmade oksüdeerumise ja nende vastastikuse mõju tõttu
Hüdrofoobses tuumas leiduvad hüdrofiilsed aminohappejäägid võivad üksteisega suhelda kasutades iooniline Ja vesiniksidemed(joonis 1.8).
Ioon- ja vesiniksidemed, samuti hüdrofoobsed vastasmõjud on nõrgad: nende energia ei ole palju suurem kui molekulide soojusliikumise energia toatemperatuuril. Valgu konformatsiooni säilitab paljude selliste nõrkade sidemete moodustumine. Kuna valku moodustavad aatomid on pidevas liikumises, on võimalik mõned nõrgad sidemed katkestada ja moodustada teisi, mis põhjustab polüpeptiidahela üksikute osade kergeid liikumisi. Seda valkude omadust muuta konformatsiooni osade katkemise ja teiste nõrkade sidemete moodustamise tulemusena nimetatakse nn. konformatsiooniline labiilsus.
Inimkehas on süsteemid, mis toetavad homöostaas- sisekeskkonna püsivus terve keha jaoks teatud vastuvõetavates piirides. Homöostaasi tingimustes ei riku väikesed konformatsiooni muutused valkude üldist struktuuri ja talitlust. Valgu funktsionaalselt aktiivset konformatsiooni nimetatakse omapärane konformatsioon. Muutused sisekeskkonnas (näiteks glükoosi, Ca ioonide, prootonite jne kontsentratsioon) toovad kaasa muutusi konformatsioonis ja valgu funktsioonide katkemist.
Mõnede valkude tertsiaarne struktuur on stabiliseerunud disulfiidsidemed, moodustub kahe jäägi -SH rühmade interaktsiooni tõttu
 Riis. 1.9. Disulfiidsideme moodustumine valgumolekulis
Riis. 1.9. Disulfiidsideme moodustumine valgumolekulis
tsüsteiin (joonis 1.9). Enamikul intratsellulaarsetel valkudel ei ole nende tertsiaarses struktuuris kovalentseid disulfiidsidemeid. Nende olemasolu on iseloomulik raku poolt sekreteeritavatele valkudele, mis tagab nende suurema stabiilsuse rakuvälistes tingimustes. Seega on disulfiidsidemed insuliini ja immunoglobuliinide molekulides.
Insuliin- valkhormoon, mis sünteesitakse kõhunäärme β-rakkudes ja eritub verre vastusena glükoosi kontsentratsiooni suurenemisele veres. Insuliini struktuuris on kaks disulfiidsidet, mis ühendavad polüpeptiidi A- ja B-ahelaid ning üks disulfiidside A-ahela sees (joonis 1.10).
 Riis. 1.10. Disulfiidsidemed insuliini struktuuris
Riis. 1.10. Disulfiidsidemed insuliini struktuuris
5. Valkude supersekundaarne struktuur. Erineva primaarse struktuuri ja funktsioonidega valkudes avastatakse neid mõnikord sekundaarsete struktuuride sarnased kombinatsioonid ja suhtelised asukohad, mida nimetatakse supersekundaarseks struktuuriks. Sellel on vahepealne positsioon sekundaarse ja tertsiaarse struktuuri vahel, kuna see on sekundaarstruktuuri elementide spetsiifiline kombinatsioon valgu tertsiaarse struktuuri moodustamisel. Supersekundaarsetel struktuuridel on spetsiifilised nimed, nagu "α-heeliks-pöörake heeliks", "leutsiintõmblukk", "tsink-sõrmed" jne. Sellised supersekundaarsed struktuurid on iseloomulikud DNA-d siduvatele valkudele.
"Leutsiini tõmblukk." Seda tüüpi supersekundaarset struktuuri kasutatakse kahe valgu ühendamiseks. Interakteeruvate valkude pinnal on α-spiraalsed piirkonnad, mis sisaldavad vähemalt nelja leutsiinijääki. Leutsiinijäägid α-heeliksis asuvad teineteisest kuue aminohappe kaugusel. Kuna α-heeliksi iga pööre sisaldab 3,6 aminohappejääki, paiknevad leutsiiniradikaalid iga teise pöörde pinnal. Ühe valgu α-heeliksi leutsiinijäägid võivad interakteeruda teise valgu leutsiinijääkidega (hüdrofoobsed interaktsioonid), ühendades need omavahel (joonis 1.11.). Paljud DNA-d siduvad valgud toimivad oligomeersetes kompleksides, kus üksikud subühikud on üksteisega ühendatud "leutsiini tõmblukkude" abil.
 Riis. 1.11. "Leutsiini tõmblukk" kahe valgu α-spiraalsete piirkondade vahel
Riis. 1.11. "Leutsiini tõmblukk" kahe valgu α-spiraalsete piirkondade vahel
Selliste valkude näide on histoonid. Histoonid- tuumavalgud, mis sisaldavad suurt hulka positiivselt laetud aminohappeid - arginiini ja lüsiini (kuni 80%). Histooni molekulid kombineeritakse kaheksat monomeeri sisaldavateks oligomeerseteks kompleksideks, kasutades "leutsiini tõmblukke", hoolimata nende molekulide olulisest homonüümsest laengust.
"Tsingi sõrm"- DNA-d siduvatele valkudele iseloomulik supersekundaarse struktuuriga variant, mis on valgu pinnal pikliku fragmendi kujul ja sisaldab umbes 20 aminohappejääki (joonis 1.12). Laiendatud sõrme kuju toetab tsingi aatom, mis on seotud nelja aminohapperadikaaliga - kahe tsüsteiini jäägi ja kahe histidiini jäägiga. Mõnel juhul on histidiini jääkide asemel tsüsteiini jäägid. Kaks tihedalt asetsevat tsüsteiinijääki eraldatakse ülejäänud kahest Gisili jäägist Cys järjestusega, mis koosneb ligikaudu 12 aminohappejäägist. See valgu piirkond moodustab α-heeliksi, mille radikaalid võivad spetsiifiliselt seonduda DNA peamise soone regulatoorsete piirkondadega. Individuaalne seondumise spetsiifilisus
 Riis. 1.12. Tsingisõrme struktuuri moodustavate DNA-d siduvate valkude piirkonna esmane struktuur (tähed tähistavad selle struktuuri moodustavaid aminohappeid)
Riis. 1.12. Tsingisõrme struktuuri moodustavate DNA-d siduvate valkude piirkonna esmane struktuur (tähed tähistavad selle struktuuri moodustavaid aminohappeid)
Reguleeriv DNA-d siduv valk sõltub tsingi sõrme piirkonnas paiknevate aminohappejääkide järjestusest. Sellised struktuurid sisaldavad eelkõige steroidhormoonide retseptoreid, mis osalevad transkriptsiooni reguleerimises (DNA-st RNA-le teabe lugemine).
TEEMA 1.2. VALGU TOIMIMISE ALUSED. VALGU FUNKTSIOONI MÕJUTAVAD ARVIVAHENDID KUI LIGANDID
1. Valgu aktiivne kese ja selle interaktsioon ligandiga. Tertsiaarse struktuuri moodustumisel moodustub funktsionaalselt aktiivse valgu pinnale tavaliselt süvendisse piirkond, mille moodustavad primaarstruktuuris üksteisest kaugel asuvad aminohapperadikaalid. Seda piirkonda, millel on antud valgu jaoks ainulaadne struktuur ja mis on võimeline spetsiifiliselt interakteeruma konkreetse molekuli või sarnaste molekulide rühmaga, nimetatakse valgu-ligandi sidumissaidiks või aktiivseks saidiks. Ligandid on molekulid, mis interakteeruvad valkudega.
Kõrge spetsiifilisus Valgu interaktsiooni ligandiga tagab aktiivtsentri struktuuri komplementaarsus ligandi struktuuriga.
Vastastikune täiendavus- see on interakteeruvate pindade ruumiline ja keemiline vastavus. Aktiivne tsenter ei pea mitte ainult ruumiliselt vastama selles sisalduvale ligandile, vaid ka sidemed (ioonsed, vesiniku- ja hüdrofoobsed interaktsioonid) peavad tekkima aktiivsesse keskusesse kuuluvate radikaalide funktsionaalrühmade ja ligandit hoidva ligandi vahel. aktiivses keskuses (joonis 1.13 ).
 Riis. 1.13. Valgu täiendav interaktsioon ligandiga
Riis. 1.13. Valgu täiendav interaktsioon ligandiga
Mõned ligandid, mis on seotud valgu aktiivse keskmega, mängivad valkude funktsioneerimisel abistavat rolli. Selliseid ligande nimetatakse kofaktoriteks ja valke, mis sisaldavad mittevalgulist osa, nimetatakse komplekssed valgud(erinevalt lihtsatest valkudest, mis koosnevad ainult valguosast). Valguga kindlalt seotud mittevalgulist osa nimetatakse proteeside rühm. Näiteks müoglobiin, hemoglobiin ja tsütokroomid sisaldavad proteesrühma, heemi, mis sisaldab raua iooni, mis on tugevalt kinnitatud aktiivse keskusega. Heemi sisaldavaid kompleksseid valke nimetatakse hemoproteiinideks.
Kui valkudele on kinnitatud spetsiifilised ligandid, avaldub nende valkude funktsioon. Seega avaldab albumiin, vereplasma kõige olulisem valk, oma transpordifunktsiooni, sidudes aktiivsesse keskusesse hüdrofoobsed ligandid, nagu rasvhapped, bilirubiin, mõned ravimid jne (joonis 1.14).
Peptiidahela kolmemõõtmelise struktuuriga interakteeruvad ligandid võivad olla mitte ainult madala molekulmassiga orgaanilised ja anorgaanilised molekulid, vaid ka makromolekulid:
DNA (eespool käsitletud DNA-d siduvate valkude näited);
polüsahhariidid;
 Riis. 1.14. Genotüübi ja fenotüübi seos
Riis. 1.14. Genotüübi ja fenotüübi seos
DNA molekulis kodeeritud inimese valkude ainulaadne primaarstruktuur realiseerub rakkudes ainulaadse konformatsiooni, aktiivse keskstruktuuri ja valgu funktsioonide kujul.
Nendel juhtudel tunneb valk ära ligandi spetsiifilise piirkonna, mis on proportsionaalne ja komplementaarne seondumiskohaga. Seega on hepatotsüütide pinnal hormooni insuliini retseptorvalgud, millel on samuti valguline struktuur. Insuliini koostoime retseptoriga põhjustab muutusi selle konformatsioonis ja signaalisüsteemide aktiveerumist, mis viib toitainete säilitamiseni hepatotsüütides pärast sööki.
Seega Valkude funktsioneerimine põhineb valgu aktiivse tsentri spetsiifilisel interaktsioonil ligandiga.
2. Domeeni struktuur ja selle roll valkude funktsioneerimisel. Globulaarsete valkude pikad polüpeptiidahelad volditakse sageli mitmeks kompaktseks, suhteliselt sõltumatuks piirkonnaks. Neil on sõltumatu tertsiaarne struktuur, mis meenutab globulaarsete valkude oma, ja neid nimetatakse domeenid. Tänu valkude domeenistruktuurile on nende tertsiaarset struktuuri lihtsam moodustada.
Domeenivalkudes paiknevad ligandi sidumissaidid sageli domeenide vahel. Seega on trüpsiin proteolüütiline ensüüm, mida toodab kõhunäärme eksokriinne osa ja mis on vajalik toiduvalkude seedimiseks. Sellel on kahedomeeniline struktuur ja trüpsiini ja selle ligandi - toiduvalgu - sidumise keskus asub kahe domeeni vahelises soones. Aktiivses keskuses luuakse tingimused, mis on vajalikud konkreetse toiduvalgu saidi efektiivseks sidumiseks ja selle peptiidsidemete hüdrolüüsiks.
Valgus olevad erinevad domeenid võivad liikuda üksteise suhtes, kui aktiivne keskus interakteerub ligandiga (joonis 1.15).
Heksokinaas- ensüüm, mis katalüüsib glükoosi fosforüülimist ATP abil. Ensüümi aktiivne sait asub kahe domeeni vahelises pilus. Kui heksokinaas seondub glükoosiga, sulguvad seda ümbritsevad domeenid ja substraat jääb lõksu, kus toimub fosforüülimine (vt joonis 1.15).
 Riis. 1.15. Heksokinaasi domeenide seondumine glükoosiga
Riis. 1.15. Heksokinaasi domeenide seondumine glükoosiga
Mõnedes valkudes täidavad domeenid erinevate ligandidega seondudes iseseisvaid funktsioone. Selliseid valke nimetatakse multifunktsionaalseteks.
3. Ravimid on ligandid, mis mõjutavad valkude talitlust. Valkude interaktsioon ligandidega on spetsiifiline. Kuid valgu ja selle aktiivse tsentri konformatsioonilise labiilsuse tõttu on võimalik valida mõni muu aine, mis võiks samuti interakteeruda valguga aktiivses keskuses või molekuli muus osas.
Loodusliku ligandiga struktuurilt sarnast ainet nimetatakse ligandi struktuurne analoog või mittelooduslik ligand. Samuti interakteerub see valguga aktiivses kohas. Ligandi struktuurne analoog võib mõlemad suurendada valgu funktsiooni (agonist), ja vähendada seda (antagonist). Ligand ja selle struktuursed analoogid konkureerivad üksteisega samas kohas valguga seondumise eest. Selliseid aineid nimetatakse konkureerivad modulaatorid valgu funktsioonide (regulaatorid). Paljud ravimid toimivad valgu inhibiitoritena. Mõned neist saadakse looduslike ligandide keemilise modifitseerimise teel. Valgu funktsioonide inhibiitorid võivad olla ravimid ja mürgid.
Atropiin on M-kolinergiliste retseptorite konkureeriv inhibiitor. Atsetüülkoliin on neurotransmitter, mis edastab närviimpulsse kolinergiliste sünapside kaudu. Ergastamise läbiviimiseks peab sünaptilisse pilusse vabanev atsetüülkoliin interakteeruma postsünaptilise membraani retseptorvalguga. Leiti kaks tüüpi kolinergilised retseptorid:
M retseptor lisaks atsetüülkoliinile interakteerub selektiivselt muskariiniga (kärbseseene toksiin). M - kolinergilised retseptorid esinevad silelihastel ja põhjustavad atsetüülkoliiniga koostoimel nende kokkutõmbumist;
H retseptor spetsiifiliselt nikotiiniga seonduv. N-kolinergilised retseptorid asuvad vöötlihaste sünapsides.
Spetsiifiline inhibiitor M-kolinergilised retseptorid on atropiin. Seda leidub belladonna ja henbane taimedes.
 Atropiinil on atsetüülkoliinile sarnase struktuuri ja ruumilise paigutusega funktsionaalsed rühmad, mistõttu on ta M-kolinergiliste retseptorite konkureeriv inhibiitor. Arvestades, et atsetüülkoliini seondumine M-kolinergiliste retseptoritega põhjustab silelihaste kontraktsiooni, kasutatakse atropiini nende spasme leevendava ravimina. (spasmoodiline). Seega on teada, et atropiini kasutatakse silmalihaste lõdvestamiseks silmapõhja vaatamisel, samuti spasmide leevendamiseks seedetrakti koolikute ajal. M-kolinergilised retseptorid esinevad ka kesknärvisüsteemis (KNS), mistõttu suured atropiiniannused võivad põhjustada kesknärvisüsteemi soovimatuid reaktsioone: motoorset ja vaimset agitatsiooni, hallutsinatsioone, krampe.
Atropiinil on atsetüülkoliinile sarnase struktuuri ja ruumilise paigutusega funktsionaalsed rühmad, mistõttu on ta M-kolinergiliste retseptorite konkureeriv inhibiitor. Arvestades, et atsetüülkoliini seondumine M-kolinergiliste retseptoritega põhjustab silelihaste kontraktsiooni, kasutatakse atropiini nende spasme leevendava ravimina. (spasmoodiline). Seega on teada, et atropiini kasutatakse silmalihaste lõdvestamiseks silmapõhja vaatamisel, samuti spasmide leevendamiseks seedetrakti koolikute ajal. M-kolinergilised retseptorid esinevad ka kesknärvisüsteemis (KNS), mistõttu suured atropiiniannused võivad põhjustada kesknärvisüsteemi soovimatuid reaktsioone: motoorset ja vaimset agitatsiooni, hallutsinatsioone, krampe.
Ditiliin on H-kolinergiliste retseptorite konkureeriv agonist, mis pärsib neuromuskulaarsete sünapside funktsiooni.
 Skeletilihaste neuromuskulaarsed sünapsid sisaldavad H-kolinergilisi retseptoreid. Nende koostoime atsetüülkoliiniga põhjustab lihaste kontraktsioone. Mõnede kirurgiliste operatsioonide, aga ka endoskoopiliste uuringute käigus kasutatakse ravimeid, mis põhjustavad skeletilihaste lõdvestamist. (lihasrelaksandid). Nende hulka kuulub ditiliin, mis on atsetüülkoliini struktuurne analoog. See kinnitub H-kolinergiliste retseptorite külge, kuid erinevalt atsetüülkoliinist hävitab seda ensüüm atsetüülkoliinesteraas väga aeglaselt. Pikaajalise ioonikanalite avanemise ja membraani püsiva depolarisatsiooni tagajärjel häiritakse närviimpulsside juhtivust ja tekib lihaste lõdvestumine. Esialgu avastati need omadused curare mürgis, mistõttu selliseid ravimeid nimetatakse curare moodi.
Skeletilihaste neuromuskulaarsed sünapsid sisaldavad H-kolinergilisi retseptoreid. Nende koostoime atsetüülkoliiniga põhjustab lihaste kontraktsioone. Mõnede kirurgiliste operatsioonide, aga ka endoskoopiliste uuringute käigus kasutatakse ravimeid, mis põhjustavad skeletilihaste lõdvestamist. (lihasrelaksandid). Nende hulka kuulub ditiliin, mis on atsetüülkoliini struktuurne analoog. See kinnitub H-kolinergiliste retseptorite külge, kuid erinevalt atsetüülkoliinist hävitab seda ensüüm atsetüülkoliinesteraas väga aeglaselt. Pikaajalise ioonikanalite avanemise ja membraani püsiva depolarisatsiooni tagajärjel häiritakse närviimpulsside juhtivust ja tekib lihaste lõdvestumine. Esialgu avastati need omadused curare mürgis, mistõttu selliseid ravimeid nimetatakse curare moodi.
TEEMA 1.3. VALGIDE DENATUREERIMINE JA NENDE SPONTAANSE RENATIVEERIMISE VÕIMALUS
1. Kuna valkude natiivne konformatsioon säilib nõrkade interaktsioonide tõttu, põhjustavad muutused valku ümbritseva keskkonna koostises ja omadustes, kokkupuude keemiliste reagentidega ja füüsikaliste teguritega nende konformatsiooni muutumist (konformatsioonilise labiilsuse omadus). Suure hulga sidemete katkemine viib natiivse konformatsiooni hävimiseni ja valkude denatureerumiseni.
Valkude denatureerimine- see on nende natiivse konformatsiooni hävitamine denatureerivate ainete mõjul, mis on põhjustatud valgu ruumilist struktuuri stabiliseerivate nõrkade sidemete katkemisest. Denaturatsiooniga kaasneb valgu ainulaadse kolmemõõtmelise struktuuri ja aktiivse keskuse hävimine ning bioloogilise aktiivsuse kadu (joonis 1.16).
Kõik ühe valgu denatureeritud molekulid omandavad juhusliku konformatsiooni, mis erineb sama valgu teistest molekulidest. Aktiivtsentrit moodustavad aminohapperadikaalid osutuvad üksteisest ruumiliselt kaugeks, s.t. valgu spetsiifiline seondumiskoht ligandiga hävib. Denatureerimise ajal jääb valkude esmane struktuur muutumatuks.
Denatureerivate ainete kasutamine bioloogilistes uuringutes ja meditsiinis. Biokeemilistes uuringutes eemaldatakse enne madala molekulmassiga ühendite määramist bioloogilises materjalis valgud tavaliselt lahusest. Sel eesmärgil kasutatakse kõige sagedamini trikloroäädikhapet (TCA). Pärast TCA lisamist lahusele denatureeritud valgud sadestuvad ja neid on lihtne filtreerimisega eemaldada (tabel 1.1).
Meditsiinis kasutatakse denatureerivaid aineid sageli meditsiiniliste instrumentide ja materjalide steriliseerimiseks autoklaavides (denatureeriv aine on kõrge temperatuur) ning antiseptikuna (alkohol, fenool, kloramiin) patogeenset mikrofloorat sisaldavate saastunud pindade töötlemisel.
2. Valkude spontaanne taasaktiveerumine- tõend valkude primaarstruktuuri, konformatsiooni ja funktsiooni determinismi kohta. Üksikud valgud on ühe geeni produktid, millel on identne aminohappejärjestus ja mis omandavad rakus sama konformatsiooni. Põhimõtteline järeldus, et valgu primaarstruktuur sisaldab juba teavet selle konformatsiooni ja funktsiooni kohta, tehti mõne valgu (eriti ribonukleaasi ja müoglobiini) võime põhjal spontaanselt renativeerida - taastada pärast denatureerimist nende natiivne konformatsioon.
Ruumiliste valgustruktuuride moodustamine toimub isekoosnemise meetodil – spontaanne protsessiga, mille käigus unikaalse primaarstruktuuriga polüpeptiidahelal on kalduvus omandada konformatsiooni, mille vaba energia lahuses on madalaim. Ensüümi ribonukleaasiga tehtud katses kirjeldati võimet renativeerida valke, mis säilitavad oma esmase struktuuri pärast denatureerimist.
Ribonukleaas on ensüüm, mis lõhub RNA molekulis üksikute nukleotiidide vahelisi sidemeid. Sellel globulaarsel valgul on üks polüpeptiidahel, mille tertsiaarne struktuur on stabiliseeritud paljude nõrkade ja nelja disulfiidsidemega.
Ribonukleaasi töötlemine uureaga, mis lõhub molekulis vesiniksidemeid, ja redutseeriva ainega, mis lõhub disulfiidsidemeid, viib ensüümi denatureerumiseni ja selle aktiivsuse kadumiseni.
Denatureerivate ainete eemaldamine dialüüsiga viib valgu konformatsiooni ja funktsiooni taastamiseni, s.t. uuestisünniks. (joonis 1.17).
 Riis. 1.17. Ribonukleaasi denatureerimine ja renatureerimine
Riis. 1.17. Ribonukleaasi denatureerimine ja renatureerimine
A - ribonukleaasi natiivne konformatsioon, mille tertsiaarstruktuuris on neli disulfiidsidet; B - denatureeritud ribonukleaasi molekul;
B - taasaktiveeritud ribonukleaasi molekul, millel on taastatud struktuur ja funktsioon
1. Täitke tabel 1.2.
Tabel 1.2. Aminohapete klassifikatsioon radikaalide polaarsuse järgi
2. Kirjutage tetrapeptiidi valem:
Asp – Pro – Fen – Liz
a) tõsta esile korduvad rühmad peptiidis, mis moodustavad peptiidi karkassi, ja varieeruvad rühmad, mida esindavad aminohapperadikaalid;
b) märgistada N- ja C-otsad;
c) tõsta esile peptiidsidemed;
d) kirjutada teine peptiid, mis koosneb samadest aminohapetest;
e) loendab sarnase aminohappe koostisega tetrapeptiidi võimalike variantide arvu.
3. Imetajate neurohüpofüüsi kahe struktuurselt sarnase ja evolutsiooniliselt lähedase peptiidhormooni – oksütotsiini ja vasopressiini – võrdleva analüüsi näitel valkude primaarstruktuuri rolli selgitamine (tabel 1.3).
Tabel 1.3. Oksütotsiini ja vasopressiini struktuur ja funktsioonid
 Selle jaoks:
Selle jaoks:
a) võrrelda kahe peptiidi koostist ja aminohappejärjestust;
b) leida kahe peptiidi primaarstruktuuri sarnasus ja nende bioloogilise toime sarnasus;
c) leida erinevusi kahe peptiidi ehituses ja nende funktsioonides;
d) teha järeldus peptiidide primaarstruktuuri mõju kohta nende funktsioonidele.
4. Kirjeldage keravalkude konformatsiooni kujunemise peamisi etappe (sekundaarsed, tertsiaarsed struktuurid, supersekundaarse struktuuri mõiste). Märkige valgustruktuuride moodustamisel osalevate sidemete tüübid. Millised aminohapperadikaalid võivad osaleda hüdrofoobsete interaktsioonide, ioonsete, vesiniksidemete moodustamisel.
Too näiteid.
5. Defineerige mõiste "valkude konformatsiooniline labiilsus", näidake selle olemasolu ja olulisuse põhjused.
6. Laiendage järgmise fraasi tähendust: "Valkude funktsioneerimine põhineb nende spetsiifilisel interaktsioonil ligandiga", kasutades termineid ja selgitades nende tähendust: valgu konformatsioon, aktiivne keskus, ligand, komplementaarsus, valgu funktsioon.
7. Selgitage ühe näite abil, mis on domeenid ja milline on nende roll valkude toimimises.
ENESEKOHTAMISÜLESANDED
1. Matš.
Funktsionaalne rühm aminohapete radikaalis:
A. Karboksüülrühm B. Hüdroksüülrühm C Guanidiinrühm D. Tioolrühm E. Aminorühm
2. Vali õiged vastused.
Polaarsete laenguta radikaalidega aminohapped on:
A. Cis B. Asn
B. Glu G. Kolm
3. Vali õiged vastused.
Aminohappe radikaalid:
A. Esitage primaarstruktuuri spetsiifilisus B. Osalege tertsiaarse struktuuri moodustamises
B. Asuvad valgu pinnal ja mõjutavad selle lahustuvust D. Moodustavad aktiivse tsentri
D. Osaleda peptiidsidemete moodustamises
4. Vali õiged vastused.
Aminohapperadikaalide vahel võivad tekkida hüdrofoobsed interaktsioonid:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Vali õiged vastused.
Aminohapperadikaalide vahel võivad tekkida ioonilised sidemed:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Gis Asp D. Asn apr
6. Vali õiged vastused.
Aminohapperadikaalide vahel võivad tekkida vesiniksidemed:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Matš.
Valgu struktuuri moodustamisel osaleva sideme tüüp:
A. Esmastruktuur B. Teisene struktuur
B. Tertsiaarne struktuur
D. Ülesekundaarne struktuur E. Konformatsioon.
1. Vesiniksidemed peptiidi karkassi aatomite vahel
2. Nõrgad sidemed aminohapperadikaalide funktsionaalrühmade vahel
3. Sidemed aminohapete α-amino- ja α-karboksüülrühmade vahel
8. Vali õiged vastused. Trüpsiin:
A. Proteolüütiline ensüüm B. Sisaldab kahte domeeni
B. Hüdrolüüsib tärklist
D. Aktiivne sait asub domeenide vahel. D. Koosneb kahest polüpeptiidahelast.
9. Vali õiged vastused. Atropiin:
A. Neurotransmitter
B. Atsetüülkoliini struktuurne analoog
B. Interakteerub H-kolinergiliste retseptoritega
D. Tugevdab närviimpulsside juhtivust kolinergiliste sünapside kaudu
D. M-kolinergiliste retseptorite konkureeriv inhibiitor
10. Valige õiged väited. Valkudes:
A. Esmastruktuur sisaldab teavet selle aktiivse saidi struktuuri kohta
B. Aktiivne keskus moodustub esmase struktuuri tasandil
B. Konformatsioon on jäigalt fikseeritud kovalentsete sidemetega
D. Aktiivne sait võib interakteeruda sarnaste ligandide rühmaga
valkude D konformatsioonilise labiilsuse tõttu. Muutused keskkonnas võivad mõjutada aktiivsete valkude afiinsust
keskus ligandiks
1. 1-B, 2-G, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
PÕHIMÕISTED JA MÕISTED
1. Valk, polüpeptiid, aminohapped
2. Primaarsed, sekundaarsed, tertsiaarsed valgustruktuurid
3. Konformatsioon, natiivse valgu konformatsioon
4. Kovalentsed ja nõrgad sidemed valkudes
5. Konformatsiooniline labiilsus
6. Valgu aktiivne sait
7. Ligandid
8. Valkude voltimine
9. Ligandide struktuursed analoogid
10. Domeeni valgud
11. Liht- ja kompleksvalgud
12. Valkude denatureerimine, denatureerivad ained
13. Valkude taasaktiveerimine
Probleeme lahendama
"Valkude struktuurne korraldus ja nende funktsioneerimise alused"
1. Valgu – hemoglobiin A (HbA) põhiülesanne on hapniku transport kudedesse. Inimpopulatsioonis tuntakse selle valgu mitmeid vorme, millel on muutunud omadused ja funktsioon – nn ebanormaalsed hemoglobiinid. Näiteks on leitud, et sirprakulise aneemia (HbS) patsientide punastes verelibledes leiduv hemoglobiin S on madala hapniku osarõhu tingimustes (nagu venoosse vere puhul) vähe lahustuv. See viib selle valgu agregaatide moodustumiseni. Valk kaotab oma funktsiooni, sadestub ja punased verelibled muutuvad ebakorrapärase kujuga (mõned neist moodustavad sirbi kuju) ja hävivad põrnas tavapärasest kiiremini. Selle tulemusena areneb sirprakuline aneemia.
Ainus erinevus HbA primaarses struktuuris leiti hemoglobiini β-ahela N-terminaalses piirkonnas. Võrrelge β-ahela N-terminaalseid piirkondi ja näidake, kuidas muutused valgu primaarstruktuuris mõjutavad selle omadusi ja funktsioone.
 Selle jaoks:
Selle jaoks:
a) kirjutage üles aminohapete valemid, mille poolest HbA erinevad, ja võrrelge nende aminohapete omadusi (polaarsus, laeng).
b) teha järeldus lahustuvuse vähenemise ja kudedesse hapniku transpordi katkemise põhjuse kohta.
2. Joonisel on kujutatud valgu struktuuri skeem, millel on ligandiga seondumiskeskus (aktiivne keskus). Selgitage, miks valk on ligandi valikul selektiivne. Selle jaoks:
a) pidage meeles, mis on valgu aktiivne kese ja arvestage joonisel näidatud valgu aktiivse tsentri struktuuriga;
b) kirjutage aktiivse keskuse moodustavate aminohapperadikaalide valemid;
c) joonistada ligand, mis võiks spetsiifiliselt interakteeruda valgu aktiivse saidiga. Märkige sellel funktsionaalsed rühmad, mis võivad moodustada sidemeid aminohappe radikaalidega, mis moodustavad aktiivse keskuse;
d) näidata sidemete tüüpe, mis tekivad aktiivse saidi ligandi ja aminohappe radikaalide vahel;
e) selgitada, millel põhineb valgu-ligandi interaktsiooni spetsiifilisus.
 3.
Joonisel on näidatud valgu ja mitme ligandi aktiivne koht.
3.
Joonisel on näidatud valgu ja mitme ligandi aktiivne koht.
Tehke kindlaks, milline ligand interakteerub kõige tõenäolisemalt valgu aktiivse saidiga ja miks.
 Mis tüüpi sidemed tekivad valgu-ligandi kompleksi moodustumisel?
Mis tüüpi sidemed tekivad valgu-ligandi kompleksi moodustumisel?
4. Looduslike valgu ligandide struktuurseid analooge saab kasutada ravimitena valkude aktiivsuse muutmiseks.
Atsetüülkoliin on neuromuskulaarsete sünapside ergastuse ülekande vahendaja. Kui atsetüülkoliin interakteerub valkudega - skeletilihaste postsünaptilise membraani retseptoritega, avanevad ioonikanalid ja toimub lihaste kokkutõmbumine. Ditiliin on ravim, mida kasutatakse teatud operatsioonide puhul lihaste lõdvestamiseks, kuna see häirib närviimpulsside ülekannet neuromuskulaarsete sünapside kaudu. Selgitage ditiliini kui lihasrelaksandina toimemehhanismi. Selle jaoks:
a) kirjutab atsetüülkoliini ja ditiliini valemid ning võrdleb nende struktuure;
b) kirjeldage ditiliini lõõgastava toime mehhanismi.
5. Mõne haiguse korral tõuseb patsiendi kehatemperatuur, mida peetakse keha kaitsvaks reaktsiooniks. Kõrge temperatuur on aga kehavalkudele kahjulik. Selgitage, miks üle 40 °C temperatuuridel valkude talitlus häirub ja tekib oht inimese elule. Selleks pidage meeles:
1) Valkude struktuur ja sidemed, mis hoiavad selle struktuuri natiivses konformatsioonis;
2) Kuidas muutub valkude struktuur ja talitlus temperatuuri tõustes?;
3) Mis on homöostaas ja miks on see inimese tervise säilitamiseks oluline.
Moodulüksus 2 OLIGOMEERILISED VALGUD KUI REGULEERIVATE MÕJUTE SIHTMÄRGID. VALKUDE STRUKTUURILINE JA FUNKTSIOONNE MITMEKESISUS. VALKUDE ERALDAMISE JA PUHASTAMISE MEETODID
Õppeeesmärgid Oskama:
1. Kasutage teadmisi oligomeersete valkude struktuuri ja funktsioonide kohta, et mõista nende funktsioonide reguleerimise adaptiivseid mehhanisme.
2. Selgitage chaperoonide rolli valkude konformatsiooni sünteesil ja säilitamisel rakutingimustes.
3. Selgitage elu ilmingute mitmekesisust organismis sünteesitavate valkude struktuuride ja funktsioonide mitmekesisuse kaudu.
4. Analüüsige valkude struktuuri ja nende funktsioonide vahelist seost, kasutades võrdlusnäiteid seotud hemoproteiinide - müoglobiini ja hemoglobiini, samuti immunoglobuliinide perekonna viie valkude klassi esindajatega.
5. Rakendada teadmisi valkude füüsikalis-keemiliste omaduste iseärasustest, et valida meetodeid nende puhastamiseks teistest valkudest ja lisanditest.
6. Kliinilise diagnoosi kinnitamiseks või täpsustamiseks tõlgendada vereplasma valkude kvantitatiivse ja kvalitatiivse koostise tulemusi.
Tea:
1. Oligomeersete valkude struktuuri tunnused ja adaptiivsed mehhanismid nende funktsioonide reguleerimiseks hemoglobiini näitel.
2. Tšaperoonide struktuur ja funktsioonid ning nende tähtsus valkude natiivse konformatsiooni säilitamisel rakutingimustes.
3. Valkude perekondadeks liitmise põhimõtted nende konformatsiooni ja funktsioonide sarnasuse alusel immunoglobuliinide näitel.
4. Valkude eraldamise meetodid nende füüsikalis-keemiliste omaduste tunnuste alusel.
5. Vereplasma elektroforees kui valkude kvalitatiivse ja kvantitatiivse koostise hindamise meetod.
TEEMA 1.4. OLIGOMEERSETE VALKUDE STRUKTUURI JA TOIMIMISE OMADUSED HEMOGLOBIINI NÄITE KOHTA
1. Paljud valgud sisaldavad mitmeid polüpeptiidahelaid. Selliseid valke nimetatakse oligomeerne, ja üksikud ketid - protomeerid. Oligomeersete valkude protomeerid on ühendatud paljude nõrkade mittekovalentsete sidemetega (hüdrofoobsed, ioonsed, vesinikud). Interaktsioon
protomeerid viiakse läbi tänu täiendavus nende kokkupuutepinnad.
Oligomeersete valkude protomeeride arv võib olla väga erinev: hemoglobiin sisaldab 4 protomeeri, ensüüm aspartaataminotransferaas 12 ja tubaka mosaiikviiruse valk sisaldab 2120 mittekovalentsete sidemetega ühendatud protomeeri. Järelikult võivad oligomeersed valgud olla väga suure molekulmassiga.
Ühe protomeeri interaktsiooni teistega võib pidada valgu-ligandi interaktsiooni erijuhtumiks, kuna iga protomeer toimib ligandina teiste protomeeride jaoks. Protomeeride arvu ja ühendamise meetodit valgus nimetatakse kvaternaarne valgu struktuur.
Valgud võivad sisaldada sama või erineva struktuuriga protomeere, näiteks homodimeerid on valgud, mis sisaldavad kahte identset protomeeri, heterodimeerid aga kahte erinevat protomeeri sisaldavad valgud.
Kui valgud sisaldavad erinevaid protomeere, siis nendele võivad moodustuda erinevate liganditega sidumiskeskused, mis erinevad struktuurilt. Kui ligand seondub aktiivse saidiga, avaldub selle valgu funktsioon. Erineval protomeeril asuvat keskust nimetatakse allosteeriliseks (erineb aktiivsest). Kontakteerumine allosteeriline ligand või efektor, see täidab reguleerivat funktsiooni (joonis 1.18). Allosteerilise tsentri interaktsioon efektoriga põhjustab konformatsioonilisi muutusi kogu oligomeerse valgu struktuuris selle konformatsioonilise labiilsuse tõttu. See mõjutab aktiivse saidi afiinsust konkreetse ligandi suhtes ja reguleerib selle valgu funktsiooni. Muutust kõigi protomeeride konformatsioonis ja funktsioonis oligomeerse valgu interaktsiooni käigus vähemalt ühe ligandiga nimetatakse kooperatiivseteks konformatsioonilisteks muutusteks. Nimetatakse efektoreid, mis suurendavad valgu funktsiooni aktivaatorid, ja efektorid, mis pärsivad selle funktsiooni - inhibiitorid.
Seega on oligomeersetel valkudel, aga ka domeenistruktuuriga valkudel, võrreldes monomeersete valkudega uus omadus – võime allosteeriliselt reguleerida funktsioone (regulatsioon valgu külge erinevate ligandide kinnitumisega). Seda saab näha kahe tihedalt seotud kompleksvalgu, müoglobiini ja hemoglobiini struktuuri ja funktsioonide võrdlemisel.
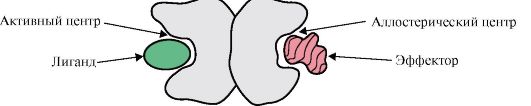 Riis. 1.18. Dimeerse valgu struktuuri skeem
Riis. 1.18. Dimeerse valgu struktuuri skeem
2. Müoglobiini ruumistruktuuride kujunemine ja funktsioneerimine.
Müoglobiin (Mb) on punastes lihastes leiduv valk, mille põhiülesanne on intensiivseks lihastööks vajalike O 2 -varude loomine. Mb on kompleksvalk, mis sisaldab valguosa – apoMb ja mittevalgulist osa – heemi. ApoMB primaarstruktuur määrab selle kompaktse kerakujulise konformatsiooni ja aktiivse tsentri struktuuri, mille külge kinnitub müoglobiini mittevalguline osa heem. Verest lihastesse tulev hapnik seondub müoglobiini Fe+ 2 heemidega. Mb on monomeerne valk, millel on väga kõrge afiinsus O 2 suhtes, seetõttu toimub hapniku vabanemine müoglobiini poolt vaid intensiivse lihastöö korral, kui O 2 osarõhk järsult langeb.
Mv konformatsiooni kujunemine. Punastes lihastes sünteesitakse translatsiooni ajal ribosoomidel MB primaarne struktuur, mida esindab 153 aminohappejäägist koosnev spetsiifiline järjestus. Mb sekundaarstruktuur sisaldab kaheksat α-heeliksit, mida nimetatakse ladina tähtedega A-st H-ni, mille vahel on mitte-spiraalsed piirkonnad. Mb tertsiaarstruktuur on kompaktse kerakujuline, mille süvendis paikneb aktiivne kese F ja E α-spiraali vahel (joon. 1.19).
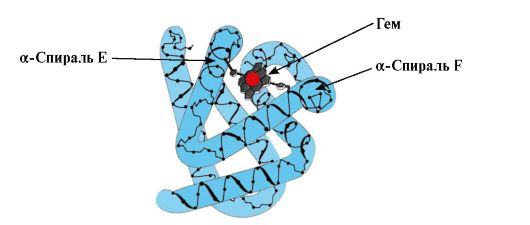 Riis. 1.19. Müoglobiini struktuur
Riis. 1.19. Müoglobiini struktuur
3. MV aktiivkeskuse struktuuri ja toimimise tunnused. Mb aktiivse tsentri moodustavad valdavalt hüdrofoobsed aminohapperadikaalid, mis on primaarstruktuuris üksteisest kaugel (näiteks Tri 3 9 ja Fen 138) Vees halvasti lahustuvad ligandid - heem ja O 2 - kinnituvad aktiivsele keskusele. Heem on apoMB spetsiifiline ligand (joonis 1.20), mille aluse moodustavad neli pürroolitsüklit, mis on ühendatud metenüülsildadega; keskel on Fe+ 2 aatom, mis on nelja koordinatsioonisidemega ühendatud pürroolitsüklite lämmastikuaatomitega. Mb aktiivses keskuses on lisaks hüdrofoobsetele aminohapperadikaalidele ka kahe aminohappe jäägid hüdrofiilsete radikaalidega - Gis E 7(Gis 64) ja GIS F 8(Tema 93) (Joon. 1.21).
 Riis. 1.20. Heemi struktuur - müoglobiini ja hemoglobiini mittevalguline osa
Riis. 1.20. Heemi struktuur - müoglobiini ja hemoglobiini mittevalguline osa
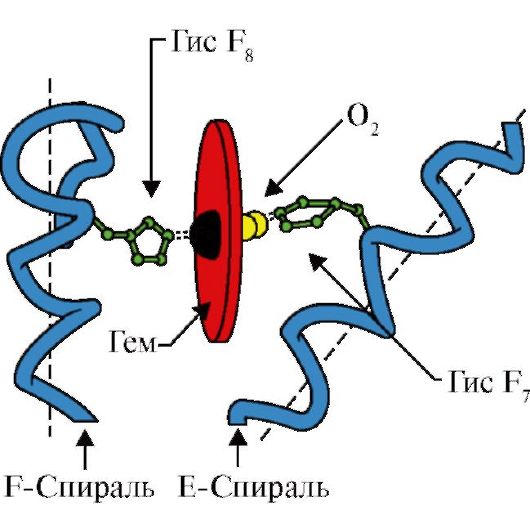 Riis. 1.21. Heemi ja O2 asukoht apomüoglobiini ja hemoglobiini protomeeride aktiivses kohas
Riis. 1.21. Heemi ja O2 asukoht apomüoglobiini ja hemoglobiini protomeeride aktiivses kohas
Heem on rauaaatomi kaudu kovalentselt seotud Tema F8-ga. O 2 kinnitub raua külge teisel pool heemtasandit. Tema E 7 on vajalik O 2 õigeks orientatsiooniks ja hõlbustab hapniku lisamist Fe + 2 heemile
GIS F 8 moodustab Fe+ 2-ga koordinatsioonisideme ja fikseerib heemi kindlalt aktiivses keskuses. Gis E 7 vajalik õigeks orienteerumiseks teise ligandi - O 2 - aktiivses keskuses selle interaktsiooni ajal Fe + 2 heemiga. Heemi mikrokeskkond loob tingimused O 2 tugevaks, kuid pöörduvaks seondumiseks Fe + 2-ga ja takistab vee sisenemist hüdrofoobsesse aktiivsesse kohta, mis võib viia selle oksüdeerumiseni Fe + 3-ks.
Mb monomeerne struktuur ja selle aktiivne tsenter määrab valgu kõrge afiinsuse O 2 suhtes.
4. Hb oligomeerne struktuur ja Hb afiinsuse reguleerimine O 2 ligandide suhtes. Inimese hemoglobiinid- valkude perekond, nagu müoglobiin, mis on seotud kompleksvalkudega (hemoproteiinidega). Need on tetrameerse struktuuriga ja sisaldavad kahte α-ahelat, kuid erinevad ülejäänud kahe polüpeptiidahela (2α-, 2x-ahelad) struktuuri poolest. Teise polüpeptiidahela struktuur määrab nende Hb vormide funktsioneerimise tunnused. Umbes 98% täiskasvanu punaliblede hemoglobiinist on hemoglobiin A(2α-, 2p-ahelad).
Loote arengu ajal toimivad kaks peamist hemoglobiini tüüpi: embrüonaalne Hb(2α, 2ε), mida leidub loote arengu varases staadiumis ja hemoglobiin F (loote)- (2α, 2γ), mis asendab varajase loote hemoglobiini emakasisese arengu kuuendal kuul ja alles pärast sündi asendatakse Hb A-ga.
HB A on valk, mis on seotud täiskasvanud inimese punastes verelibledes leiduva müoglobiiniga (MB). Selle üksikute protomeeride struktuur on sarnane müoglobiini omaga. Müoglobiini ja hemoglobiini protomeeride sekundaarne ja tertsiaarne struktuur on väga sarnased, hoolimata asjaolust, et nende polüpeptiidahelate primaarstruktuuris on identsed ainult 24 aminohappejääki (hemoglobiini protomeeride sekundaarstruktuur, nagu ka müoglobiin, sisaldab kaheksat α-heeliksit, tähistatakse ladina tähtedega vahemikus A kuni H ja tertsiaarne struktuur on kompaktse kera kujuline). Kuid erinevalt müoglobiinist on hemoglobiinil oligomeerne struktuur, mis koosneb neljast mittekovalentsete sidemetega ühendatud polüpeptiidahelast (joonis 1.22).
Iga Hb protomeer on seotud mittevalgulise osaga – heemi ja naaberprotomeeridega. Hb valguosa seos heemiga on sarnane müoglobiini omaga: valgu aktiivses keskuses on heemi hüdrofoobsed osad ümbritsetud hüdrofoobsete aminohapperadikaalidega, välja arvatud His F 8 ja His E 7, mis paiknevad mõlemal pool heemitasandit ja mängivad sarnast rolli valgu toimimises ja hapnikuga seondumises (vt müoglobiini struktuur).
 Riis. 1.22. Hemoglobiini oligomeerne struktuur
Riis. 1.22. Hemoglobiini oligomeerne struktuur
Pealegi, Gis E 7 täidab olulist lisaroll Nv toimimises. Vaba heemi afiinsus CO suhtes on 25 000 korda kõrgem kui O2 suhtes. CO moodustub kehas väikestes kogustes ja arvestades selle suurt afiinsust heemi suhtes, võib see häirida rakkude eluks vajaliku O 2 transporti. Kuid hemoglobiini koostises ületab heemi afiinsus süsinikmonooksiidi suhtes ainult 200 korda afiinsust O 2 suhtes, kuna aktiivses keskuses on His E 7. Ülejäänud osa sellest aminohappest loob optimaalsed tingimused heemi sidumiseks O 2 -ga ja nõrgendab heemi koostoimet COga.
5. HB põhiülesanne on O2 transportimine kopsudest kudedesse. Erinevalt monomeersest müoglobiinist, millel on väga kõrge afiinsus O2 suhtes ja mis täidab hapniku säilitamise funktsiooni punastes lihastes, pakub hemoglobiini oligomeerne struktuur:
1) HB kiire küllastumine hapnikuga kopsudes;
2) HB võime vabastada kudedes hapnikku suhteliselt kõrge O 2 osarõhul (20-40 mm Hg);
3) võimalus reguleerida Hb afiinsust O 2 suhtes.
6. Kooperatiivsed muutused hemoglobiini protomeeride konformatsioonis kiirendavad O 2 seondumist kopsudes ja selle vabanemist kudedesse. Kopsudes soodustab O 2 kõrge osarõhk selle seondumist Hb-ga nelja protomeeri (2α ja 2β) aktiivses kohas. Iga protomeeri aktiivne kese, nagu ka müoglobiinil, asub kahe α-heeliksi (F ja E) vahel hüdrofoobses taskus. See sisaldab mittevalgulist osa – heemi, mis on seotud valguosaga paljude nõrkade hüdrofoobsete interaktsioonide kaudu ning üht tugevat sidet Fe 2 + heemi ja His F 8 vahel (vt joonis 1.21).
Tänu sellele sidemele His F 8-ga ulatub desoksühemoglobiinis Fe 2 + aatom heemi tasapinnast histidiini poole. O 2 seondumine Fe 2 +-ga toimub heemi teisel küljel His E 7 piirkonnas, kasutades ühtainsat vaba koordinatsiooni sidet. Tema E 7 loob optimaalsed tingimused O 2 seondumiseks heemi rauaga.
O 2 lisamine ühe protomeeri Fe + 2 aatomile põhjustab selle liikumise heemi tasapinnale, millele järgneb sellega seotud histidiini jääk
 Riis. 1.23. Hemoglobiini protomeeri konformatsiooni muutus kombineerimisel O2-ga
Riis. 1.23. Hemoglobiini protomeeri konformatsiooni muutus kombineerimisel O2-ga
See toob kaasa muutuse kõigi polüpeptiidahelate konformatsioonis nende konformatsioonilise labiilsuse tõttu. Teiste ahelate konformatsiooni muutmine hõlbustab nende interaktsiooni järgnevate O 2 molekulidega.
Neljas O 2 molekul seondub hemoglobiiniga 300 korda kergemini kui esimene (joonis 1.24).
 Riis. 1.24. Kooperatiivsed muutused hemoglobiini protomeeride konformatsioonis selle interaktsiooni ajal O2-ga
Riis. 1.24. Kooperatiivsed muutused hemoglobiini protomeeride konformatsioonis selle interaktsiooni ajal O2-ga
Kudedes lõhustub iga järgnev O 2 molekul kergemini kui eelmine, seda ka protomeeride konformatsiooni kooperatiivsete muutuste tõttu.
7. Orgaaniliste ainete katabolismi käigus tekkivad CO 2 ja H+ vähendavad hemoglobiini afiinsust O 2 suhtes võrdeliselt nende kontsentratsiooniga. Rakkude funktsioneerimiseks vajalik energia toodetakse peamiselt mitokondrites orgaaniliste ainete oksüdatsiooni käigus, kasutades hemoglobiiniga kopsudest tarnitud O 2. Orgaaniliste ainete oksüdatsiooni tulemusena tekivad nende lagunemise lõppsaadused: CO 2 ja K 2 O, mille hulk on võrdeline käimasolevate oksüdatsiooniprotsesside intensiivsusega.
CO 2 difundeerub rakkudest verre ja tungib punastesse verelibledesse, kus ensüümi karbanhüdraasi toimel muundub see süsihappeks. See nõrk hape dissotsieerub prootoniks ja vesinikkarbonaadi iooniks.
H+ on võimelised ühinema Tema radikaalidega 14 6 hemoglobiini α- ja β-ahelates, st. heemist kaugemal asuvates piirkondades. Hemoglobiini protoneerimine vähendab selle afiinsust O 2 suhtes, soodustab O 2 eemaldamist oksüHb-st, desoksüHb moodustumist ja suurendab kudede varustatust hapnikuga võrdeliselt moodustunud prootonite arvuga (joonis 1.25).
Vabanenud hapniku hulga suurenemist sõltuvalt H+ kontsentratsiooni suurenemisest punastes verelibledes nimetatakse Bohri efektiks (nimetatud Taani füsioloogi Christian Bohri järgi, kes selle efekti esmakordselt avastas).
Kopsudes soodustab hapniku kõrge osarõhk selle seondumist deoksüHb-ga, mis vähendab valgu afiinsust H + suhtes. Süsihappe toimel vabanevad prootonid reageerivad vesinikkarbonaatidega, moodustades CO 2 ja H 2 O

 Riis. 1.25. Hb afiinsuse sõltuvus O 2 suhtes CO 2 ja prootonite kontsentratsioonist (Bohri efekt):
Riis. 1.25. Hb afiinsuse sõltuvus O 2 suhtes CO 2 ja prootonite kontsentratsioonist (Bohri efekt):
A- CO 2 ja H+ kontsentratsiooni mõju O 2 vabanemisele kompleksist HB-ga (Bohri efekt); B- desoksühemoglobiini hapnikuga varustamine kopsudes, CO 2 moodustumine ja vabanemine.
Saadud CO 2 siseneb alveolaarruumi ja eemaldatakse väljahingatavas õhus. Seega reguleerivad hemoglobiini poolt kudedes vabaneva hapniku hulka orgaaniliste ainete katabolismi produktid: mida intensiivsem on ainete lagunemine, näiteks füüsilise koormuse ajal, seda suurem on CO 2 ja H + kontsentratsioon ning seda rohkem hapnikku. kuded saavad Hb afiinsuse vähenemise tulemusena O 2 suhtes.
8. Hb afiinsuse allosteeriline reguleerimine O2 suhtes ligandi - 2,3-bisfosfoglütseraadi poolt. Erütrotsüütides sünteesitakse hemoglobiini allosteeriline ligand 2,3-bisfosfoglütseraat (2,3-BPG) glükoosi oksüdatsiooniproduktist - 1,3-bisfosfoglütseraadist. Normaalsetes tingimustes on 2,3-BPG kontsentratsioon kõrge ja võrreldav Hb kontsentratsiooniga. 2,3-BPG-l on tugev negatiivne laeng -5.
 Bisfosfoglütseraat kudede kapillaarides, seondudes desoksühemoglobiiniga, suurendab hapniku vabanemist kudedesse, vähendades Hb afiinsust O 2 suhtes.
Bisfosfoglütseraat kudede kapillaarides, seondudes desoksühemoglobiiniga, suurendab hapniku vabanemist kudedesse, vähendades Hb afiinsust O 2 suhtes.
Tetrameerse hemoglobiini molekuli keskel on õõnsus. Selle moodustavad kõigi nelja protomeeri aminohappejäägid (vt joonis 1.22). Kudede kapillaarides põhjustab Hb protoneerimine (Bohri efekt) heemi raua ja O2 vahelise sideme katkemise. Molekulis
desoksühemoglobiin, võrreldes oksühemoglobiiniga tekivad täiendavad ioonsidemed, mis ühendavad protomeere, mille tulemusena suurenevad tsentraalse õõnsuse mõõtmed võrreldes oksühemoglobiiniga. Keskne õõnsus on koht, kus 2,3-BPG seondub hemoglobiiniga. Keskse õõnsuse suuruse erinevuse tõttu saab 2,3-BPG kinnituda ainult desoksühemoglobiini külge.
2,3-BPG interakteerub hemoglobiiniga kohas, mis on eemal valgu aktiivsetest keskustest ja kuulub allosteeriline(regulatiivsed) ligandid ja Hb keskne õõnsus on allosteeriline keskus. 2,3-BPG-l on tugev negatiivne laeng ja see interakteerub Hb kahe β-ahela viie positiivselt laetud rühmaga: Val N-terminaalse α-aminorühma ja Lys 82 His 143 radikaalidega (joonis 1.26).
 Riis. 1.26. BPG desoksühemoglobiini keskses õõnes
Riis. 1.26. BPG desoksühemoglobiini keskses õõnes
BPG seondub igas β-ahelas kolme positiivselt laetud rühmaga.
Kudede kapillaarides interakteerub tekkiv desoksühemoglobiin 2,3-BPG-ga ning β-ahelate positiivselt laetud radikaalide ja negatiivselt laetud ligandi vahel tekivad ioonilised sidemed, mis muudavad valgu konformatsiooni ja vähendavad Hb afiinsust O2 suhtes. . Hb afiinsuse vähenemine O 2 suhtes soodustab O 2 tõhusamat vabanemist koesse.
Kopsudes interakteerub kõrge osarõhu korral hapnik Hb-ga, liitudes heemi rauaga; sel juhul muutub valgu konformatsioon, tsentraalne õõnsus väheneb ja 2,3-BPG nihkub allosteerilisest keskusest välja
Seega on oligomeersetel valkudel monomeersete valkudega võrreldes uued omadused. Ligandide kinnitamine kohtadesse
üksteisest ruumiliselt kaugel (allosteerilised), võivad põhjustada konformatsioonilisi muutusi kogu valgumolekulis. Tänu interaktsioonile regulatoorsete ligandidega toimub konformatsiooni muutus ja valgu molekuli funktsiooni kohanemine keskkonnamuutustega.
TEEMA 1.5. VALGIDE NATIIVSE KONFORMAATSIOONI SÄILITAMINE RAKU TINGIMUSTEL
Rakkudes polüpeptiidahelate sünteesil, nende transportimisel läbi membraanide raku vastavatesse osadesse, voltimise protsessis (natiivse konformatsiooni moodustumise) ja oligomeersete valkude kokkupanemise käigus, samuti nende funktsioneerimisel, vaheühend. , valgu struktuuris tekivad agregatsioonile kalduvad ebastabiilsed konformatsioonid. Hüdrofoobsed radikaalid, mis on tavaliselt natiivses konformatsioonis valgu molekuli sees peidetud, ilmuvad pinnale ebastabiilse konformatsioonina ja kipuvad ühinema teiste vees halvasti lahustuvate valkude rühmadega. Kõikide teadaolevate organismide rakkudes on leitud spetsiaalsed valgud, mis tagavad rakuvalkude optimaalse voltimise, stabiliseerivad nende natiivset konformatsiooni funktsioneerimisel ning mis kõige olulisem – säilitavad homöostaasi häirete korral rakusiseste valkude struktuuri ja funktsioonid. Neid valke nimetatakse "saatjad" mis tähendab prantsuse keeles "lapsehoidja".
1. Molekulaarsed šaperoonid ja nende roll valkude denaturatsiooni ennetamisel.
Chaperonid (CH) klassifitseeritakse nende allüksuste massi järgi. Kõrge molekulmassiga chaperonide mass on 60 kuni 110 kDa. Nende hulgas on enim uuritud kolme klassi: Sh-60, Sh-70 ja Sh-90. Iga klass sisaldab seotud valkude perekonda. Seega sisaldab Sh-70 valke molekulmassiga 66 kuni 78 kDa. Madala molekulmassiga chaperonide molekulmass on 40 kuni 15 kDa.
Saatjate hulgas on konstitutiivne valgud, mille kõrge basaalsüntees ei sõltu stressi mõjust keharakkudele ja indutseeritav, mille süntees tavatingimustes on nõrk, kuid stressi korral suureneb järsult. Indutseeritavaid chaperone nimetatakse ka "kuumašoki valkudeks", kuna need avastati esmakordselt kõrge temperatuuriga kokku puutunud rakkudes. Rakkudes on valkude kõrge kontsentratsiooni tõttu osaliselt denatureeritud valkude spontaanne taasaktiveerimine raskendatud. Sh-70 võib ära hoida denaturatsiooni algust ja aidata taastada valkude loomulikku konformatsiooni. Molekulaarsed chaperonid-70- väga konserveerunud valkude klass, mida leidub raku kõigis osades: tsütoplasmas, tuumas, endoplasmaatilises retikulumis, mitokondrites. Ühe polüpeptiidahela Ш-70 karboksüülotsas on piirkond, mis on soon, mis on võimeline interakteeruma pikkade peptiididega
7 kuni 9 aminohappejääki, mis on rikastatud hüdrofoobsete radikaalidega. Sellised piirkonnad globulaarsetes valkudes esinevad ligikaudu iga 16 aminohappe järel. Sh-70 on võimeline kaitsma valke temperatuuri inaktiveerimise eest ning taastama osaliselt denatureeritud valkude konformatsiooni ja aktiivsust.
2. Tšaperoonide roll valkude voltimisel. Valgusünteesi käigus ribosoomil sünteesitakse polüpeptiidi N-terminaalne piirkond enne C-terminaalset piirkonda. Natiivse konformatsiooni moodustamiseks on vaja valgu täielikku aminohappejärjestust. Valgusünteesi käigus suudavad chaperones-70 oma aktiivse tsentri struktuuri tõttu sulgeda polüpeptiidi piirkondi, mis on altid agregatsioonile ja on rikastatud hüdrofoobsete aminohapperadikaalidega, kuni süntees on lõppenud (joonis 1.27, A).
 Riis. 1.27. Tšaperonide osalemine valkude voltimises
Riis. 1.27. Tšaperonide osalemine valkude voltimises
A - chaperons-70 osalemine hüdrofoobsete interaktsioonide vältimises sünteesitud polüpeptiidi lõikude vahel; B - valgu natiivse konformatsiooni moodustumine chaperone kompleksis
Paljud kõrgmolekulaarsed valgud, millel on keeruline konformatsioon, näiteks domeeni struktuur, voldivad Sh-60 moodustatud spetsiaalses ruumis. Ш-60 toimib oligomeerse kompleksina, mis koosneb 14 subühikust. Need moodustavad kaks õõnsat rõngast, millest igaüks koosneb seitsmest allüksusest, need rõngad on omavahel ühendatud. Iga Sh-60 alaühik koosneb kolmest domeenist: apikaalne (apikaalne), rikastatud hüdrofoobsete radikaalidega, mis on suunatud rõnga õõnsusele, vahepealne ja ekvatoriaalne (joonis 1.28).
 Riis. 1.28. Tšaperoniini kompleksi struktuur, mis koosneb 14 Ш-60
Riis. 1.28. Tšaperoniini kompleksi struktuur, mis koosneb 14 Ш-60
A - külgvaade; B - pealtvaade
Sünteesitud valgud, mille pinnal on voldimata molekulidele iseloomulikke elemente, eriti hüdrofoobseid radikaale, sisenevad chaperooni rõngaste õõnsustesse. Nende õõnsuste spetsiifilises keskkonnas otsitakse võimalikke konformatsioone, kuni leitakse ainus, mis on energeetiliselt soodsaim (joon. 1.27, B). Konformatsioonide moodustumise ja valgu vabanemisega kaasneb ekvatoriaalpiirkonnas ATP hüdrolüüs. Tavaliselt nõuab selline chaperonist sõltuv voltimine märkimisväärsel hulgal energiat.
Lisaks osalemisele valkude kolmemõõtmelise struktuuri moodustamises ja osaliselt denatureeritud valkude renativeerimises on šaperoonid vajalikud ka selliste fundamentaalsete protsesside toimumiseks nagu oligomeersete valkude kokkupanek, denatureeritud valkude äratundmine ja transportimine lüsosoomidesse, valkude transport läbi membraanide ja osalemine valgukomplekside aktiivsuse reguleerimises.
TEEMA 1.6. VALGUDE MITMESUGUS. VALGUPERED: IMMUNOGLOBULIINIDE NÄIDE
1. Valgud mängivad üksikute rakkude ja kogu hulkrakulise organismi elus määravat rolli ning nende funktsioonid on üllatavalt mitmekesised. Selle määravad ära valkude primaarstruktuuri ja konformatsioonide omadused, aktiivse keskuse ainulaadne struktuur ja võime siduda spetsiifilisi ligande.
Vaid väga väike osa kõigist võimalikest peptiidahelate variantidest võib omandada stabiilse ruumilise struktuuri; enamus
neist võib olla palju konformatsioone ligikaudu sama Gibbsi energiaga, kuid erinevate omadustega. Enamike teadaolevate valkude esmane struktuur, mis on valitud bioloogilise evolutsiooni teel, tagab ühe konformatsiooni erakordse stabiilsuse, mis määrab selle valgu funktsioneerimise omadused.
2. Valgupered. Sama bioloogilise liigi sees võivad aminohappejääkide asendused viia erinevate valkude tekkeni, mis täidavad seotud funktsioone ja millel on homoloogsed aminohappejärjestused. Sellistel sugulasvalkudel on silmatorkavalt sarnased konformatsioonid: α-heeliksite ja/või β-struktuuride arv ja suhteline asend, enamik polüpeptiidahelate pöördeid ja painutusi on sarnased või identsed. Polüpeptiidahela homoloogsete piirkondadega, sarnase konformatsiooni ja seotud funktsioonidega valgud liigitatakse valgu perekondadesse. Valguperekondade näited: seriini proteinaasid, immunoglobuliinide perekond, müoglobiini perekond.
Seriini proteinaasid- proteolüütiliste ensüümide funktsiooni täitev valkude perekond. Nende hulka kuuluvad seedeensüümid – kümotrüpsiin, trüpsiin, elastaas ja paljud vere hüübimisfaktorid. Nendel valkudel on 40% positsioonidest identsed aminohapped ja väga sarnane konformatsioon (joonis 1.29).

Riis. 1.29. Elastaasi (A) ja kümotrüpsiini (B) ruumilised struktuurid
Mõned aminohapete asendused on põhjustanud muutusi nende valkude substraadi spetsiifilisuses ja funktsionaalse mitmekesisuse tekkimist perekonna sees.
3. Immunoglobuliinide perekond. Immuunsüsteemi toimimises mängivad tohutut rolli immunoglobuliinide superperekonna valgud, mis hõlmavad kolme valgu perekonda:
Antikehad (immunoglobuliinid);
T-lümfotsüütide retseptorid;
Peamise histocompatibility kompleksi valgud – MHC klassid 1 ja 2 (Major Histocompatibility Complex).
Kõik need valgud on domeenistruktuuriga, koosnevad homoloogsetest immuunlaadsetest domeenidest ja täidavad sarnaseid funktsioone: interakteeruvad võõrstruktuuridega, mis on lahustunud veres, lümfis või rakkudevahelises vedelikus (antikehad) või paiknevad rakkude pinnal (oma või välismaa).
4. Antikehad- spetsiifilised valgud, mida toodavad B-lümfotsüüdid vastusena võõrstruktuuri sisenemisele organismi, nn antigeen.
Antikehade struktuuri tunnused
Kõige lihtsamad antikehamolekulid koosnevad neljast polüpeptiidahelast: kaks identset kerget - L, mis sisaldavad umbes 220 aminohapet, ja kaks identset rasket - H, mis koosnevad 440-700 aminohappest. Kõik neli ahelat antikeha molekulis on ühendatud paljude mittekovalentsete sidemetega ja nelja disulfiidsidemega (joonis 1.30).
Antikeha kerged ahelad koosnevad kahest domeenist: varieeruvast domeenist (VL), mis asub polüpeptiidahela N-terminaalses piirkonnas, ja konstantsest domeenist (CL), mis asub C-otsas. Rasketel ahelatel on tavaliselt neli domeeni: üks muutuja (VH), mis asub N-otsas, ja kolm konstantset domeeni (CH1, CH2, CH3) (vt joonis 1.30). Igal immunoglobuliini domeenil on β-lehe pealisehitus, milles kaks tsüsteiinijääki on seotud disulfiidsidemega.
Kahe konstantse domeeni CH1 ja CH2 vahel on piirkond, mis sisaldab suurt hulka proliinijääke, mis takistavad selles segmendis sekundaarse struktuuri teket ja naaber-H-ahelate interaktsiooni. See liigendpiirkond annab antikehamolekulile paindlikkuse. Raske ja kerge ahela varieeruvate domeenide vahel on kaks identset antigeeni siduvat saiti (aktiivsed saidid antigeenide sidumiseks), seetõttu nimetatakse selliseid antikehi sageli nn. kahevalentsed. Antigeeni seondumises antikehaga ei osale mitte kogu mõlema ahela varieeruvate piirkondade aminohappejärjestus, vaid ainult 20-30 aminohapet, mis paiknevad iga ahela hüpervarieeruvates piirkondades. Just need piirkonnad määravad igat tüüpi antikehade ainulaadse võime interakteeruda vastava komplementaarse antigeeniga.
Antikehad on üks keha kaitseliine sissetungivate võõrorganismide vastu. Nende toimimise võib jagada kaheks etapiks: esimene etapp on antigeeni äratundmine ja sidumine võõrorganismide pinnal, mis on võimalik tänu antigeeni sidumiskohtade olemasolule antikeha struktuuris; teine etapp on antigeeni inaktiveerimise ja hävitamise protsessi käivitamine. Teise etapi spetsiifilisus sõltub antikehade klassist. Raskeid ahelaid on viis klassi, mis erinevad üksteisest konstantsete domeenide struktuuri poolest: α, δ, ε, γ ja μ, mille järgi eristatakse viit immunoglobuliinide klassi: A, D, E, G ja M.
Raskete ahelate struktuursed omadused annavad raskete ahelate liigendpiirkondadele ja C-terminaalsetele piirkondadele igale klassile iseloomuliku konformatsiooni. Pärast antigeeni seondumist antikehaga määravad konformatsioonilised muutused konstantsetes domeenides antigeeni eemaldamise tee.
 Riis. 1. 30. IgG domeenistruktuur
Riis. 1. 30. IgG domeenistruktuur
Immunoglobuliinid M
Immunoglobuliinidel M on kaks vormi.
Monomeerne vorm- B-lümfotsüütide arenemise teel toodetud 1. klassi antikehad. Seejärel lülituvad paljud B-rakud üle teiste klasside antikehade tootmisele, kuid sama antigeeni sidumissaidiga. IgM on põimitud membraani ja toimib antigeeni äratundmise retseptorina. IgM-i integreerumine rakumembraani on võimalik tänu 25 hüdrofoobse aminohappejäägi olemasolule piirkonna sabaosas.
IgM sekretoorne vorm sisaldab viit monomeerset subühikut, mis on omavahel seotud disulfiidsidemetega ja täiendavat polüpeptiid-J ahelat (joonis 1.31). Selle vormi monomeeride rasked ahelad ei sisalda hüdrofoobset saba. Pentameeril on 10 antigeeni sidumissaiti ja seetõttu on see efektiivne esimesena kehasse siseneva antigeeni äratundmisel ja eemaldamisel. IgM sekretoorne vorm on primaarse immuunvastuse käigus verre erituvate antikehade peamine klass. IgM-i seondumine antigeeniga muudab IgM-i konformatsiooni ja indutseerib selle seondumist komplemendisüsteemi esimese valgukomponendiga (komplemendisüsteem on valkude kogum, mis osaleb antigeeni hävitamises) ja selle süsteemi aktiveerimise. Kui antigeen asub mikroorganismi pinnal, põhjustab komplemendi süsteem rakumembraani terviklikkuse häireid ja bakteriraku surma.
Immunoglobuliinid G
Kvantitatiivselt on see immunoglobuliinide klass veres ülekaalus (75% kõigist Ig-dest). IgG – monomeerid, sekundaarse immuunvastuse käigus verre erituvate antikehade põhiklass. Pärast IgG interaktsiooni mikroorganismide pinnaantigeenidega on antigeen-antikeha kompleks võimeline siduma ja aktiveerima komplementsüsteemi valke või interakteeruma makrofaagide ja neutrofiilide spetsiifiliste retseptoritega. Koostoime fagotsüütidega viib
 Riis. 1.31. IgM sekretoorse vormi struktuur
Riis. 1.31. IgM sekretoorse vormi struktuur
antigeen-antikeha komplekside imendumisele ja nende hävitamisele raku fagosoomides. IgG on ainus antikehade klass, mis on võimeline tungima läbi platsentaarbarjääri ja tagama loote emakasisese kaitse infektsioonide eest.
Immunoglobuliinid A
Peamine antikehade klass sekretsioonides (piim, sülg, hingamisteede ja soolte sekretsioon). IgA sekreteeritakse peamiselt dimeersel kujul, kus monomeerid on omavahel seotud täiendava J-ahela kaudu (joonis 1.32).
IgA ei interakteeru komplemendisüsteemi ja fagotsüütrakkudega, kuid seondudes mikroorganismidega takistavad antikehad nende kinnitumist epiteelirakkudele ja tungimist organismi.
Immunoglobuliinid E
Immunoglobuliine E esindavad monomeerid, milles rasked ε-ahelad sisaldavad sarnaselt immunoglobuliinide M μ-ahelatele ühte muutuvat ja nelja konstantset domeeni. Pärast sekretsiooni seondub IgE omaga
 Riis. 1.32. IgA struktuur
Riis. 1.32. IgA struktuur
C-terminaalsed piirkonnad vastavate retseptoritega nuumrakkude ja basofiilide pinnal. Selle tulemusena muutuvad nad nende rakkude pinnal olevate antigeenide retseptoriteks (joonis 1.33).
 Riis. 1.33. IgE interaktsioon antigeeniga nuumraku pinnal
Riis. 1.33. IgE interaktsioon antigeeniga nuumraku pinnal
Pärast antigeeni kinnitumist IgE vastavate antigeeni siduvate saitide külge saavad rakud signaali bioloogiliselt aktiivsete ainete (histamiini, serotoniini) sekreteerimiseks, mis on suures osas vastutavad põletikulise reaktsiooni tekke ja allergiliste reaktsioonide ilmnemise eest, nt. astma, urtikaaria, heinapalavik.
Immunoglobuliinid D
Immunoglobuliine D leidub seerumis väga väikestes kogustes, need on monomeerid. Rasketel δ ahelatel on üks muutuv ja kolm konstantset domeeni. IgD-d toimivad B-lümfotsüütide retseptoritena, teised funktsioonid on siiani teadmata. Spetsiifiliste antigeenide interaktsioon B-lümfotsüütide (IgD) pinnal olevate retseptoritega viib nende signaalide edastamiseni rakku ja mehhanismide aktiveerumiseni, mis tagavad antud lümfotsüütide klooni proliferatsiooni.
TEEMA 1.7. VALGIDE FÜÜSIKALISED JA KEEMILISED OMADUSED NING NENDE LAHETAMISE MEETODID
1. Üksikud valgud erinevad oma füüsikalis-keemiliste omaduste poolest:
Molekulide kuju;
Molekulmass;
Kogulaeng, mille suurus sõltub aminohapete anioonsete ja katioonsete rühmade suhtest;
Polaarsete ja mittepolaarsete aminohapperadikaalide suhe molekulide pinnal;
Vastupidavus erinevatele denatureerivatele ainetele.
2. Valkude lahustuvus sõltubülalloetletud valkude omaduste kohta, samuti söötme koostise kohta, milles valk on lahustunud (pH väärtused, soola koostis, temperatuur, muude orgaaniliste ainete olemasolu, mis võivad valguga suhelda). Valgumolekulide laengu hulk on üks nende lahustuvust mõjutavatest teguritest. Kui isoelektrilise punkti laeng kaob, agregeeruvad ja sadestuvad valgud kergemini. See on eriti tüüpiline denatureeritud valkude puhul, mille pinnale ilmuvad hüdrofoobsed aminohapperadikaalid.
Valgu molekuli pinnal on nii positiivselt kui ka negatiivselt laetud aminohapperadikaale. Nende rühmade arv ja seega ka valkude kogulaeng sõltub söötme pH-st, s.o. H+ - ja OH - rühmade kontsentratsioonide suhe. Happelises keskkonnas H+ kontsentratsiooni tõus toob kaasa karboksüülrühmade -COO - + H+ > -COOH dissotsiatsiooni pärssimise ja valkude negatiivse laengu vähenemise. Aluselises keskkonnas põhjustab OH liia sidumine prootonitega, mis moodustuvad aminorühmade -NH 3 + + OH - - NH 2 + H 2 O dissotsiatsioonil vee moodustumisega, valkude positiivse laengu vähenemise. . Nimetatakse pH väärtust, mille juures valgu puhas nulllaeng on isoelektriline punkt (IEP). IET-s on positiivselt ja negatiivselt laetud rühmade arv sama, s.t. valk on isoelektrilises olekus.
3. Üksikute valkude eraldamine. Keha struktuuri ja toimimise tunnused sõltuvad selles sünteesitud valkude komplektist. Valkude struktuuri ja omaduste uurimine on võimatu ilma neid rakust eraldamata ja teistest valkudest ja orgaanilistest molekulidest puhastamata. Üksikute valkude eraldamise ja puhastamise etapid:
Rakkude hävitamine uuritav kude ja homogenaadi saamine.
Homogenaadi eraldamine fraktsioonideks tsentrifuugimise teel, saades soovitud valku sisaldava tuuma, mitokondriaalse, tsütosoolse või muu fraktsiooni.
Selektiivne termiline denatureerimine- valgulahuse lühiajaline kuumutamine, mille käigus saab eemaldada osa denatureeritud valgulisanditest (kui valk on suhteliselt kuumakindel).
Väljasoolamine. Erinevad valgud sadestuvad lahuses erineva soolakontsentratsiooniga. Soola kontsentratsiooni järk-järgult suurendades on võimalik saada mitmeid eraldi fraktsioone, millest ühes on ülekaalus isoleeritud valgu sisaldus. Ammooniumsulfaati kasutatakse kõige sagedamini valkude fraktsioneerimiseks. Väikseima lahustuvusega valgud sadestuvad madalate soolakontsentratsioonide korral.
Geelfiltreerimine- meetod molekulide sõelumiseks läbi paisunud Sephadexi graanulite (kolmemõõtmelised pooridega dekstraani polüsahhariidahelad). Valkude läbimise kiirus Sephadexiga täidetud kolonnist sõltub nende molekulmassist: mida väiksem on valgumolekulide mass, seda kergemini need graanulitesse tungivad ja seal kauem püsivad, seda kiiremini nad sealt elueeruvad veerg.
Ultratsentrifuugimine- meetod, mis hõlmab valkude asetamist tsentrifuugitorus ultratsentrifuugi rootorisse. Kui rootor pöörleb, on valkude settimiskiirus võrdeline nende molekulmassiga: raskemate valkude fraktsioonid asuvad katseklaasi põhjale lähemal, kergemad - pinnale lähemal.
Elektroforees- meetod, mis põhineb valkude liikumiskiiruse erinevustel elektriväljas. See väärtus on võrdeline valkude laenguga. Valkude elektroforees viiakse läbi paberil (sel juhul on valkude liikumise kiirus võrdeline ainult nende laenguga) või teatud pooride suurusega polüakrüülamiidgeelis (valgu liikumise kiirus on võrdeline nende laengu ja molekulmassiga) .
Ioonivahetuskromatograafia- fraktsioneerimismeetod, mis põhineb valkude ioniseeritud rühmade sidumisel ioonvahetusvaikude (lahustumatud polümeersed materjalid) vastaslaenguga rühmadega. Valgu vaiguga seondumise tugevus on võrdeline valgu laenguga. Ioonivahetuspolümeerile adsorbeerunud valke saab maha pesta NaCl lahuste kasvavate kontsentratsioonidega; mida madalam on valgu laeng, seda väiksem on NaCl kontsentratsioon, mis on vajalik vaigu ioonrühmadega seotud valgu mahapesemiseks.
Afiinsuskromatograafia- kõige spetsiifilisem meetod üksikute valkude eraldamiseks. Valgu ligand on kovalentselt seotud inertse polümeeriga. Kui valgulahus lastakse läbi kolonni polümeeriga, adsorbeerub kolonnile ainult antud ligandile spetsiifiline valk valgu komplementaarse seondumise tõttu ligandiga.
Dialüüs- meetod, mida kasutatakse madala molekulmassiga ühendite eemaldamiseks eraldatud valgu lahusest. Meetod põhineb valkude võimetusel läbida poolläbilaskvat membraani, erinevalt madala molekulmassiga ainetest. Seda kasutatakse valkude puhastamiseks madala molekulmassiga lisanditest, näiteks sooladest pärast väljasoolamist.
KAVAVÄLISTE TÖÖDE ÜLESANDED
1. Täitke tabel. 1.4.
Tabel 1.4. Seotud valkude - müoglobiini ja hemoglobiini - struktuuri ja funktsioonide võrdlev analüüs
a) mäleta Mb ja Hb aktiivtsentri struktuuri. Millist rolli mängivad hüdrofoobsed aminohapperadikaalid nende valkude aktiivsete keskuste moodustamisel? Kirjeldage Mb ja Hb aktiivtsentri ehitust ja ligandide sellega kinnitumise mehhanisme. Millist rolli mängivad His F 8 ja His E 7 jäägid Mv iHv aktiivkeskuse toimimises?
b) millised uued omadused on monomeerse müoglobiiniga võrreldes tihedalt seotud oligomeerse valgu hemoglobiiniga? Selgitage kooperatiivsete muutuste rolli hemoglobiini molekuli protomeeride konformatsioonis, CO 2 ja prootoni kontsentratsioonide mõju hemoglobiini afiinsusele hapniku suhtes, samuti 2,3-BPG rolli Hb funktsiooni allosteerilises regulatsioonis. .
2. Iseloomustage molekulaarseid chaperone, pöörates tähelepanu nende struktuuri ja funktsiooni vahelisele seosele.
3. Millised valgud on rühmitatud perekondadesse? Tuvastage immunoglobuliinide perekonna näitel selle perekonna valkude sarnased struktuursed tunnused ja seotud funktsioonid.
4. Biokeemilistel ja meditsiinilistel eesmärkidel on sageli vaja puhastatud üksikuid valke. Selgitage, millistel valkude füüsikalis-keemilistel omadustel põhinevad nende eraldamiseks ja puhastamiseks kasutatavad meetodid.
ENESEKOHTAMISÜLESANDED
1. Vali õiged vastused.
Hemoglobiini funktsioonid:
A. O 2 transport kopsudest kudedesse B. H + transport kudedest kopsudesse
B. Konstantse vere pH hoidmine D. CO 2 transport kopsudest kudedesse
D. CO 2 transport kudedest kopsudesse
2. Vali õiged vastused. Ligandα -protomeeri Hb on: A. Heme
B. Hapnik
B. CO G. 2,3-BPG
D. β-protomeer
3. Vali õiged vastused.
Hemoglobiin erinevalt müoglobiinist:
A. on kvaternaarse struktuuriga
B. Sekundaarstruktuuri esindavad ainult α-heeliksid
B. Kuulub kompleksvalkudesse
D. Interakteerub allosteerilise ligandiga D. Seotud kovalentselt heemiga
4. Vali õiged vastused.
Hb afiinsus O2 suhtes väheneb:
A. Kui lisatakse üks O 2 molekul B. Kui üks O 2 molekul on eemaldatud
B. 2,3-BPG-ga suhtlemisel
D. Kui kinnitub protomeeridele H + D. Kui 2,3-BPG kontsentratsioon väheneb
5. Matš.
HB tüüpidele on iseloomulik:
A. Deoksüvormis moodustab see fibrillaarseid agregaate B. Sisaldab kahte α- ja kahte δ-ahelat
B. Hb domineeriv vorm täiskasvanud erütrotsüütides D. Sisaldab heemi, mille aktiivses keskuses on Fe+ 3
D. Sisaldab kahte α- ja kahte γ-ahelat 1. HbA 2.
6. Matš.
Hb ligandid:
A. Seondub allosteerilises keskuses Hb-ga
B. Sellel on väga kõrge afiinsus Hb aktiivse saidi suhtes
B. Liitudes suurendab see Hb afiinsust O 2 G suhtes. Oksüdeerib Fe+ 2 kuni Fe+ 3
D. Moodustab kovalentse sideme hisF8-ga
7. Vali õiged vastused.
Saatjad:
A. Raku kõikides osades esinevad valgud
B. Süntees suureneb stressi all
B. Osaleda denatureeritud valkude hüdrolüüsis
D. Osaleda valkude natiivse konformatsiooni säilitamises
D. Loo organellid, milles moodustub valkude konformatsioon
8. Matš. Immunoglobuliinid:
A. Sekretoorne vorm on pentameerne.
B. Ig klass, mis tungib läbi platsentaarbarjääri
B. Ig – nuumrakkude retseptor
D. Peamine Ig klass, mis esineb epiteelirakkude eritistes. D. B-lümfotsüütide retseptor, mille aktiveerimine tagab rakkude proliferatsiooni
9. Vali õiged vastused.
Immunoglobuliinid E:
A. Makrofaagide poolt toodetud B. Neil on rasked ε-ahelad.
B. Manustatud T-lümfotsüütide membraani
D. Toimivad nuumrakkude ja basofiilide membraani antigeeni retseptoritena
D. Vastutab allergiliste reaktsioonide eest
10. Vali õiged vastused.
Valkude eraldamise meetod põhineb nende molekulmassi erinevustel:
A. Geelfiltreerimine
B. Ultratsentrifuugimine
B. Polüakrüülamiidgeelelektroforees D. Ioonivahetuskromatograafia
D. Afiinsuskromatograafia
11. Vali õige vastus.
Valkude eraldamise meetod põhineb nende vees lahustuvuse erinevustel:
A. Geelfiltreerimine B. Väljasoolamine
B. Ioonivahetuskromatograafia D. Afiinsuskromatograafia
D. Elektroforees polüakrüülamiidgeelis
„ENESEKOHTAMISÜLESANDE“ VASTUSTE STANDARDID
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-G
6. 1-B, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-B
PÕHIMÕISTED JA MÕISTED
1. Oligomeersed valgud, protomeer, valkude kvaternaarne struktuur
2. Kooperatiivsed muutused protomeeride konformatsioonis
3. Bohri efekt
4. Valkude funktsioonide allosteeriline regulatsioon, allosteeriline tsenter ja allosteeriline efektor
5. Molekulaarsed chaperonid, kuumašoki valgud
6. Valguperekonnad (seriiniproteaasid, immunoglobuliinid)
7. IgM-, G-, E-, A-struktuuri ja funktsiooni seos
8. Valkude kogulaeng, valkude isoelektriline punkt
9. Elektroforees
10. Väljasoolamine
11. Geelfiltreerimine
12. Ioonivahetuskromatograafia
13. Ultratsentrifuugimine
14. Afiinsuskromatograafia
15. Vereplasma valkude elektroforees
ÜLESANDED KLASSITÖÖKS
1. Võrrelge hemoglobiini (Hb) ja müoglobiini (Mb) küllastusastmete sõltuvust hapnikuga selle osarõhust kudedes
 Riis. 1.34. Küllastumise sõltuvus Mv jaNHhapnikku selle osarõhust
Riis. 1.34. Küllastumise sõltuvus Mv jaNHhapnikku selle osarõhust
Pange tähele, et valgu hapnikuküllastuse kõverate kuju on erinev: müoglobiini puhul - hüperbool, hemoglobiini puhul - sigmoidne kuju.
1. Võrrelge hapniku osarõhu väärtusi, mille juures Mb ja Hb on O 2 -ga küllastunud 50% võrra. Millisel neist valkudest on suurem afiinsus O 2 suhtes?
2. Millised Mb struktuuriomadused määravad selle kõrge afiinsuse O 2 suhtes?
3. Millised HB struktuuriomadused võimaldavad tal vabastada O2 puhkeaega kudede kapillaarides (suhteliselt kõrge O2 osarõhu juures) ja järsult tõsta seda vabanemist töötavates lihastes? Milline oligomeersete valkude omadus annab selle efekti?
4. Arvutage, kui palju O 2 (%) hapnikuga küllastunud hemoglobiini annab puhkavatele ja töötavatele lihastele?
5. teha järeldusi valgu ehituse ja funktsiooni vahelise seose kohta.
2. Hemoglobiini poolt kapillaarides vabaneva hapniku hulk sõltub kudedes toimuvate kataboolsete protsesside intensiivsusest (Bohri efekt). Kuidas reguleerivad muutused kudede ainevahetuses Hb afiinsust O2 suhtes? CO 2 ja H+ mõju Hb afiinsusele O 2 suhtes
 1. kirjeldage Bohri efekti.
1. kirjeldage Bohri efekti.
2. mis suunas skeemil näidatud protsess kulgeb:
a) kopsude kapillaarides;
b) kudede kapillaarides?
3. Mis on Bohri efekti füsioloogiline tähtsus?
4. Miks muudab Hb interaktsioon H+-ga heemist kaugemates kohtades valgu afiinsust O 2 suhtes?
3. Hb afiinsus O2 suhtes sõltub selle ligandi kontsentratsioonist - 2,3-bisfosfoglütseraadist, mis on Hb afiinsuse allosteeriline regulaator O2 suhtes. Miks mõjutab ligandi interaktsioon aktiivsest saidist kaugel asuvas kohas valgu funktsiooni? Kuidas 2,3-BPG reguleerib Hb afiinsust O2 suhtes? Probleemi lahendamiseks vastake järgmistele küsimustele:
1. kus ja millest sünteesitakse 2,3-bisfosfoglütseraati (2,3-BPG)? Kirjutage selle valem, märkige selle molekuli laeng.
2. Millise hemoglobiini vormiga (oksü või desoksü) BPG interakteerub ja miks? Millises Hb molekuli osas interaktsioon toimub?
3. mis suunas toimub diagrammil näidatud protsess?
a) kudede kapillaarides;
b) kopsukapillaarides?
4. kus kompleksi kontsentratsioon peaks olema suurem
Nv-2,3-BFG:
a) puhkeolekus olevate lihaste kapillaarides,
b) töötavate lihaste kapillaarides (eeldusel, et erütrotsüütides on sama BPG kontsentratsioon)?
5. Kuidas muutub HB afiinsus hapniku suhtes inimese kohanemisel kõrgmäestikutingimustega, kui BPG kontsentratsioon erütrotsüütides suureneb? Mis on selle nähtuse füsioloogiline tähtsus?
4. 2,3-BPG hävimine konserveeritud vere säilitamise ajal kahjustab HB funktsioone. Kuidas muutub HB afiinsus O 2 suhtes säilinud veres, kui 2,3-BPG kontsentratsioon erütrotsüütides võib langeda 8-lt 0,5 mmol/l-le. Kas sellist verd on võimalik raskelt haigetele patsientidele üle kanda, kui 2,3-BPG kontsentratsioon taastub mitte varem kui kolme päeva pärast? Kas 2,3-BPG lisamisega verre on võimalik punaste vereliblede funktsioone taastada?
5. Pidage meeles lihtsaimate immunoglobuliinimolekulide struktuuri. Millist rolli mängivad immunoglobuliinid immuunsüsteemis? Miks nimetatakse Igsid sageli kahevalentseteks? Kuidas on Ig-ide struktuur seotud nende funktsiooniga? (Kirjeldage immunoglobuliinide klassi näite abil.)
Valkude füüsikalis-keemilised omadused ja nende eraldamise meetodid.
6. Kuidas mõjutab valgu netolaeng selle lahustuvust?
a) määrake peptiidi kogulaeng pH 7 juures
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) kuidas muutub selle peptiidi laeng pH >7, pH juures<7, рН <<7?
c) mis on valgu isoelektriline punkt (IEP) ja millises keskkonnas see asub?
Selle peptiidi IET?
d) millise pH väärtuse juures täheldatakse selle peptiidi madalaimat lahustuvust.
7. Miks hapupiim erinevalt värskest piimast keetmisel "kalgendub" (s.t. piimavalk kaseiin sadestub)? Värskes piimas on kaseiini molekulidel negatiivne laeng.
8. Üksikute valkude eraldamiseks kasutatakse geelfiltreerimist. Segu, mis sisaldas valke A, B, C molekulmassiga vastavalt 160 000, 80 000 ja 60 000, analüüsiti geelfiltratsiooniga (joonis 1.35). Paisunud geeligraanulid on läbilaskvad valkudele, mille molekulmass on alla 70 000. Mis põhimõte on selle eraldamismeetodi aluseks? Milline graafik kajastab fraktsioneerimise tulemusi õigesti? Märkige valkude A, B ja C kolonnist vabanemise järjekord.
 Riis. 1.35. Geelfiltratsiooni kasutamine valkude eraldamiseks
Riis. 1.35. Geelfiltratsiooni kasutamine valkude eraldamiseks
9. Joonisel fig. 1.36, A näitab terve inimese vereseerumi valkude paberil elektroforeesi diagrammi. Selle meetodi abil saadud valgufraktsioonide suhtelised kogused on järgmised: albumiinid 54-58%, α1-globuliinid 6-7%, α2-globuliinid 8-9%, β-globuliinid 13%, y-globuliinid 11-12%.
 Riis. 1.36 Terve inimese (A) ja patsiendi (B) vereplasma valkude elektroforees paberil
Riis. 1.36 Terve inimese (A) ja patsiendi (B) vereplasma valkude elektroforees paberil
I - y-globuliinid; II - β-globuliinid; III -α 2 -globuliin; IV -α 2 -globuliin; V - albumiinid
Paljude haigustega kaasnevad kvantitatiivsed muutused seerumi valkude koostises (düsproteineemia). Diagnoosi tegemisel ning haiguse tõsiduse ja staadiumi hindamisel võetakse arvesse nende muutuste olemust.
Oravad- suure molekulmassiga orgaanilised ühendid, mis koosnevad α-aminohappe jääkidest.
IN valgu koostis hõlmab süsinikku, vesinikku, lämmastikku, hapnikku, väävlit. Mõned valgud moodustavad komplekse teiste fosforit, rauda, tsinki ja vaske sisaldavate molekulidega.
Valkudel on suur molekulmass: munaalbumiin - 36 000, hemoglobiin - 152 000, müosiin - 500 000 Võrdluseks: alkoholi molekulmass on 46, äädikhape - 60, benseen - 78.
Valkude aminohappeline koostis
Oravad- mitteperioodilised polümeerid, mille monomeerid on α-aminohapped. Tavaliselt nimetatakse valgu monomeerideks 20 tüüpi α-aminohappeid, kuigi üle 170 neist leidub rakkudes ja kudedes.
Sõltuvalt sellest, kas aminohappeid saab inimeste ja teiste loomade kehas sünteesida, eristatakse neid: mitteasendatavad aminohapped- saab sünteesida; asendamatud aminohapped- ei saa sünteesida. Asendamatud aminohapped tuleb organismi varustada toiduga. Taimed sünteesivad igat tüüpi aminohappeid.
Sõltuvalt aminohapete koostisest valgud on: täielikud- sisaldab kogu aminohapete komplekti; defektne- nende koostises puuduvad mõned aminohapped. Kui valgud koosnevad ainult aminohapetest, nimetatakse neid lihtne. Kui valgud sisaldavad lisaks aminohapetele ka mitteaminohappelist komponenti (proteesirühma), nimetatakse neid nn. keeruline. Proteesirühma võivad esindada metallid (metalloproteiinid), süsivesikud (glükoproteiinid), lipiidid (lipoproteiinid), nukleiinhapped (nukleoproteiinid).
Kõik aminohapped sisaldavad: 1) karboksüülrühm (-COOH), 2) aminorühm (-NH2), 3) radikaal või R-rühm (ülejäänud molekul). Radikaali struktuur on erinevat tüüpi aminohapete puhul erinev. Sõltuvalt aminohapete koostises sisalduvate aminorühmade ja karboksüülrühmade arvust eristatakse neid: neutraalsed aminohapped millel on üks karboksüülrühm ja üks aminorühm; aluselised aminohapped millel on rohkem kui üks aminorühm; happelised aminohapped millel on rohkem kui üks karboksüülrühm.
Aminohapped on amfoteersed ühendid, kuna lahuses võivad nad toimida nii hapete kui alustena. Vesilahustes esinevad aminohapped erinevates ioonsetes vormides.
Peptiidside
Peptiidid- orgaanilised ained, mis koosnevad peptiidsidemetega ühendatud aminohappejääkidest.
Peptiidide moodustumine toimub aminohapete kondensatsioonireaktsiooni tulemusena. Kui ühe aminohappe aminorühm interakteerub teise aminohappe karboksüülrühmaga, tekib nende vahel kovalentne lämmastik-süsinik side, mida nimetatakse nn. peptiid. Sõltuvalt peptiidis sisalduvate aminohappejääkide arvust on olemas dipeptiidid, tripeptiidid, tetrapeptiidid jne. Peptiidsideme moodustumist võib korrata mitu korda. See viib moodustumiseni polüpeptiidid. Peptiidi ühes otsas on vaba aminorühm (nn N-ots) ja teises on vaba karboksüülrühm (nn C-ots).

Valgumolekulide ruumiline korraldus
Teatud spetsiifiliste funktsioonide täitmine valkude poolt sõltub nende molekulide ruumilisest konfiguratsioonist, lisaks on rakule energeetiliselt ebasoodne hoida valke voldimata kujul, ahela kujul, mistõttu polüpeptiidahelad läbivad voltimise, omandades ahela; teatud kolmemõõtmeline struktuur või konformatsioon. Seal on 4 taset valkude ruumiline korraldus.
Valgu esmane struktuur- aminohappejääkide paigutuse järjestus valgu molekuli moodustavas polüpeptiidahelas. Aminohapete vaheline side on peptiidside.

Kui valgumolekul koosneb vaid 10 aminohappejäägist, siis teoreetiliselt võimalike aminohapete vaheldumise järjekorra poolest erinevate valgumolekulide variantide arv on 10 20. Omades 20 aminohapet, saate neist teha veelgi mitmekesisemaid kombinatsioone. Inimese organismist on leitud umbes kümme tuhat erinevat valku, mis erinevad nii üksteisest kui ka teiste organismide valkudest.
See on valgumolekuli esmane struktuur, mis määrab valgumolekulide omadused ja selle ruumilise konfiguratsiooni. Ainuüksi ühe aminohappe asendamine teisega polüpeptiidahelas viib valgu omaduste ja funktsioonide muutumiseni. Näiteks kuuenda glutamiinaminohappe asendamine valiiniga hemoglobiini β-subühikus toob kaasa asjaolu, et hemoglobiini molekul tervikuna ei suuda täita oma põhifunktsiooni - hapniku transporti; Sellistel juhtudel tekib inimesel haigus, mida nimetatakse sirprakuliseks aneemiaks.
Sekundaarne struktuur- polüpeptiidahela järjestatud voltimine spiraaliks (näeb välja nagu pikendatud vedru). Heeliksi pöördeid tugevdavad vesiniksidemed, mis tekivad karboksüülrühmade ja aminorühmade vahel. Peaaegu kõik CO ja NH rühmad osalevad vesiniksidemete moodustamises. Need on nõrgemad kui peptiidsed, kuid palju kordi korrates annavad sellele konfiguratsioonile stabiilsuse ja jäikuse. Sekundaarse struktuuri tasemel on valgud: fibroiin (siid, ämblikuvõrk), keratiin (juuksed, küüned), kollageen (kõõlused).



Tertsiaarne struktuur- polüpeptiidahelate pakkimine gloobulitesse, mis tulenevad keemiliste sidemete (vesinik-, ioon-, disulfiid) moodustumisest ja hüdrofoobsete interaktsioonide loomisest aminohappejääkide radikaalide vahel. Peamist rolli tertsiaarse struktuuri moodustamisel mängivad hüdrofiilsed-hüdrofoobsed interaktsioonid. Vesilahustes kipuvad hüdrofoobsed radikaalid vee eest peitu pugema, koondudes gloobuli sisse, hüdrofiilsed radikaalid aga hüdratatsiooni (koostoime vee dipoolidega) tulemusena molekuli pinnale. Mõnedes valkudes stabiliseerivad tertsiaarset struktuuri kahe tsüsteiinijäägi väävliaatomite vahel moodustunud kovalentsed disulfiidsidemed. Tertsiaarse struktuuri tasemel on ensüümid, antikehad ja mõned hormoonid.

Kvaternaarne struktuur iseloomulik kompleksvalkudele, mille molekulid on moodustatud kahest või enamast gloobulist. Subühikuid hoiavad molekulis ioonsed, hüdrofoobsed ja elektrostaatilised interaktsioonid. Mõnikord tekivad kvaternaarse struktuuri moodustumisel subühikute vahel disulfiidsidemed. Enim uuritud kvaternaarse struktuuriga valk on hemoglobiini. See moodustub kahest α-subühikust (141 aminohappejääki) ja kahest β-subühikust (146 aminohappejääki). Iga alaühikuga on seotud rauda sisaldav heemimolekul.
Kui valkude ruumiline konformatsioon mingil põhjusel normaalsest hälbib, ei saa valk oma funktsioone täita. Näiteks "hullu lehma tõve" (spongiformse entsefalopaatia) põhjuseks on prioonide, närvirakkude pinnavalkude, ebanormaalne konformatsioon.
Valkude omadused
Valgumolekuli aminohappeline koostis ja struktuur määrab selle omadused. Valgud ühendavad aluselised ja happelised omadused, mille määravad aminohapperadikaalid: mida rohkem happelisi aminohappeid on valgus, seda rohkem väljenduvad selle happelised omadused. Määratakse annetamise ja H + lisamise võime valkude puhverdavad omadused; Üks võimsamaid puhvreid on punastes verelibledes sisalduv hemoglobiin, mis hoiab vere pH konstantsel tasemel. On lahustuvad valgud (fibrinogeen) ja on lahustumatud valgud, mis täidavad mehaanilisi funktsioone (fibroiin, keratiin, kollageen). On valke, mis on keemiliselt aktiivsed (ensüümid), on keemiliselt inaktiivseid valke, mis on vastupidavad erinevatele keskkonnatingimustele ja need, mis on äärmiselt ebastabiilsed.
Välised tegurid (kuumus, ultraviolettkiirgus, raskmetallid ja nende soolad, pH muutused, kiirgus, dehüdratsioon)
võib põhjustada häireid valgumolekuli struktuuris. Antud valgu molekulile omase kolmemõõtmelise konformatsiooni kadumise protsessi nimetatakse denatureerimine. Denaturatsiooni põhjuseks on teatud valgu struktuuri stabiliseerivate sidemete katkemine. Esialgu katkevad nõrgemad sidemed ja tingimuste karmistudes katkevad veelgi tugevamad. Seetõttu kaovad esmalt kvaternaar, seejärel tertsiaar- ja sekundaarstruktuurid. Ruumilise konfiguratsiooni muutus toob kaasa valgu omaduste muutumise ja selle tulemusena muudab valgu võimatuks oma loomulike bioloogiliste funktsioonide täitmise. Kui denaturatsiooniga ei kaasne primaarstruktuuri hävimine, võib see nii olla pööratav, sel juhul toimub valgule iseloomuliku konformatsiooni isetaastumine. Näiteks membraani retseptori valgud läbivad sellise denaturatsiooni. Valgu struktuuri taastamise protsessi pärast denatureerimist nimetatakse renaturatsioon. Kui valgu ruumilise konfiguratsiooni taastamine on võimatu, nimetatakse denaturatsiooni pöördumatu.
Valkude funktsioonid
| Funktsioon | Näited ja selgitused |
|---|---|
| Ehitus | Valgud osalevad rakuliste ja rakuväliste struktuuride moodustamises: need on osa rakumembraanidest (lipoproteiinid, glükoproteiinid), juustest (keratiin), kõõlustest (kollageen) jne. |
| Transport | Verevalk hemoglobiin seob hapnikku ja transpordib selle kopsudest kõikidesse kudedesse ja organitesse ning viib sealt edasi süsihappegaasi kopsudesse; Rakumembraanide koostis sisaldab spetsiaalseid valke, mis tagavad teatud ainete ja ioonide aktiivse ja rangelt selektiivse ülekande rakust väliskeskkonda ja tagasi. |
| Reguleerivad | Valguhormoonid osalevad ainevahetusprotsesside reguleerimises. Näiteks hormoon insuliin reguleerib vere glükoosisisaldust, soodustab glükogeeni sünteesi ja suurendab rasvade moodustumist süsivesikutest. |
| Kaitsev | Vastuseks võõrvalkude või mikroorganismide (antigeenide) tungimisele organismi moodustuvad spetsiaalsed valgud – antikehad, mis suudavad neid siduda ja neutraliseerida. Fibrinogeenist moodustunud fibriin aitab peatada verejooksu. |
| Mootor | Kokkutõmbuvad valgud aktiin ja müosiin tagavad mitmerakuliste loomade lihaste kontraktsiooni. |
| Signaal | Raku pinnamembraani sisse on ehitatud valgumolekulid, mis on võimelised keskkonnateguritele reageerides muutma oma tertsiaarset struktuuri, saades seeläbi väliskeskkonnast signaale ja edastades rakule käsklusi. |
| Säilitamine | Loomade kehas valke reeglina ei säilitata, välja arvatud munaalbumiin ja piimakaseiin. Kuid tänu valkudele saavad mõned ained organismis talletuda, näiteks hemoglobiini lagunemisel rauda kehast ei eemaldata, vaid see ladestub, moodustades valgu ferritiiniga kompleksi. |
| Energia | Kui 1 g valku laguneb lõpptoodeteks, vabaneb 17,6 kJ. Esiteks lagunevad valgud aminohapeteks ja seejärel lõpptoodeteks - veeks, süsinikdioksiidiks ja ammoniaagiks. Valke kasutatakse aga energiaallikana alles siis, kui muud allikad (süsivesikud ja rasvad) on ära kasutatud. |
| Katalüütiline | Valkude üks tähtsamaid funktsioone. Toetavad valgud - ensüümid, mis kiirendavad rakkudes toimuvaid biokeemilisi reaktsioone. Näiteks ribuloosbifosfaatkarboksülaas katalüüsib CO 2 fikseerimist fotosünteesi käigus. |
Ensüümid
Ensüümid, või ensüümid, on spetsiaalne valkude klass, mis on bioloogilised katalüsaatorid. Tänu ensüümidele toimuvad biokeemilised reaktsioonid tohutu kiirusega. Ensümaatiliste reaktsioonide kiirus on kümneid tuhandeid kordi (ja mõnikord miljoneid) suurem kui anorgaaniliste katalüsaatorite osalusel toimuvate reaktsioonide kiirus. Aine, millel ensüüm toimib, nimetatakse substraat.
Ensüümid on globulaarsed valgud, struktuursed omadused Ensüümid võib jagada kahte rühma: lihtsad ja keerulised. Lihtsad ensüümid on lihtsad valgud, st. koosnevad ainult aminohapetest. Komplekssed ensüümid on kompleksvalgud, st. Lisaks valguosale sisaldavad need rühma mittevalgulist laadi - kofaktor. Mõned ensüümid kasutavad kofaktoritena vitamiine. Ensüüm molekul sisaldab spetsiaalset osa, mida nimetatakse aktiivseks keskuseks. Aktiivne keskus- ensüümi väike osa (kolm kuni kaksteist aminohappejääki), kus substraadi või substraatide seondumine toimub ensüümi-substraadi kompleksi moodustamiseks. Reaktsiooni lõppedes laguneb ensüümi-substraadi kompleks ensüümiks ja reaktsiooniprodukti(de)ks. Mõnedel ensüümidel on (välja arvatud aktiivsed) allosteerilised keskused- alad, millele on kinnitatud ensüümi kiiruse regulaatorid ( allosteerilised ensüümid).

Ensümaatilise katalüüsi reaktsioone iseloomustavad: 1) kõrge efektiivsus, 2) range selektiivsus ja toimesuund, 3) substraadi spetsiifilisus, 4) peen ja täpne reguleerimine. Ensümaatilise katalüüsi reaktsioonide substraadi ja reaktsiooni spetsiifilisust selgitavad E. Fischeri (1890) ja D. Koshlandi (1959) hüpoteesid.
E. Fisher (klahviluku hüpotees) tegi ettepaneku, et ensüümi ja substraadi aktiivse saidi ruumilised konfiguratsioonid peavad üksteisele täpselt vastama. Substraati võrreldakse “võtmega”, ensüümi “lukuga”.
D. Koshland (käsikinda hüpotees) tegi ettepaneku, et substraadi struktuuri ja ensüümi aktiivse tsentri vaheline ruumiline vastavus luuakse ainult nende üksteisega suhtlemise hetkel. Seda hüpoteesi nimetatakse ka indutseeritud vastavuse hüpotees.
Ensümaatiliste reaktsioonide kiirus sõltub: 1) temperatuurist, 2) ensüümi kontsentratsioonist, 3) substraadi kontsentratsioonist, 4) pH-st. Tuleb rõhutada, et kuna ensüümid on valgud, on nende aktiivsus kõrgeim füsioloogiliselt normaalsetes tingimustes.
Enamik ensüüme saab töötada ainult temperatuurivahemikus 0–40 °C. Nendes piirides suureneb reaktsioonikiirus ligikaudu 2 korda iga 10 °C temperatuuri tõusuga. Temperatuuril üle 40 °C valk denatureerub ja ensüümide aktiivsus väheneb. Külmumislähedasel temperatuuril ensüümid inaktiveeritakse.
Substraadi koguse suurenedes suureneb ensümaatilise reaktsiooni kiirus, kuni substraadi molekulide arv võrdub ensüümi molekulide arvuga. Substraadi koguse edasise suurenemisega kiirus ei suurene, kuna ensüümi aktiivsed keskused on küllastunud. Ensüümide kontsentratsiooni suurenemine suurendab katalüütilist aktiivsust, kuna ajaühikus muundub suurem arv substraadi molekule.

Iga ensüümi jaoks on optimaalne pH väärtus, mille juures see avaldab maksimaalset aktiivsust (pepsiin - 2,0, sülje amülaas - 6,8, pankrease lipaas - 9,0). Kõrgemate või madalamate pH väärtuste korral ensüümi aktiivsus väheneb. PH järskude muutustega ensüüm denatureerub.
Allosteeriliste ensüümide kiirust reguleerivad ained, mis kinnituvad allosteeriliste tsentrite külge. Kui need ained kiirendavad reaktsiooni, nimetatakse neid aktivaatorid, kui nad aeglustavad - inhibiitorid.
Ensüümide klassifikatsioon
Vastavalt nende katalüüsitavate keemiliste muundumiste tüübile jagatakse ensüümid 6 klassi:
- oksireduktaasid(vesiniku, hapniku või elektroni aatomite ülekandmine ühelt ainelt teisele - dehüdrogenaas),
- transferaasid(metüül-, atsüül-, fosfaat- või aminorühma ülekandmine ühelt ainelt teisele - transaminaas),
- hüdrolaasid(hüdrolüüsireaktsioonid, mille käigus substraadist moodustuvad kaks toodet - amülaas, lipaas),
- lüaasid(mittehüdrolüütiline lisamine substraadile või aatomirühma eemaldamine sellest, mille puhul C-C, C-N, C-O, C-S sidemed võivad katkeda - dekarboksülaas),
- isomeraasid(molekulaarne ümberkorraldus - isomeraas),
- ligaasid(kahe molekuli ühendus C-C, C-N, C-O, C-S sidemete moodustumise tulemusena - süntetaas).
Klassid jagunevad omakorda alamklassideks ja alamklassideks. Praeguses rahvusvahelises klassifikatsioonis on igal ensüümil konkreetne kood, mis koosneb neljast punktidega eraldatud numbrist. Esimene number on klass, teine on alamklass, kolmas on alamklass, neljas on selle alaklassi ensüümi seerianumber, näiteks arginaasi kood on 3.5.3.1.
Minema loengud nr 2"Süsivesikute ja lipiidide struktuur ja funktsioonid"
Minema loengud nr 4"ATP nukleiinhapete struktuur ja funktsioonid"