Looduses leidub baktereid, millel on ainulaadne omadus eoseid moodustada. Lugeja saab artiklit lugedes teada, kuidas see protsess toimub.
Vaidlused
Eosbakterid on vastupidavamad külmumisele, kuivamisele, pikaajalisele või lühiajalisele keetmisele ja kemikaalidega kokkupuutele. On olemas bakterite moodustumise ja eoste vegetatiivsed vormid. Viimaste näited: selliste haiguste patogeenid nagu siberi katk, botulism, teetanus ja teatud tüüpi saprofüütsed mullaasukad, mida võib leida sõnnikus.
Idanemine
Kui eoste kest satub talle soodsasse keskkonda, hakkab see paisuma. See protsess jätkub, kuni kest on täielikult hävinud. Ajal, mil membraani kude puruneb, siseneb noor rakk selle suure lõhkemise kaudu väliskeskkonda.
Nii idaneb eos aeroobsetes bakterites. Anaeroobsed bakterid ei kaota sporulatsiooni käigus oma välist rakumembraani. Eos ei puutu kokku väliskeskkonnaga, tema kokkupuude toimub rakumembraaniga. Soodsate tingimuste tekkimisel satuvad toitained rakku läbi katte. Eos hakkab idanema.
Bakterid ja tooted
Teatud toodete töötlemise ja ladustamise ajal on bakterite sporulatsioon ebasoovitav. Kui see protsess toimub, on mikroorganismidega raske võidelda. Näiteks konservide spooride hävitamiseks tuleb toode steriliseerida, mis vähendab oluliselt selle kvaliteeti. Piima pikaajaliseks säilimiseks see steriliseeritakse ning see toob kaasa selle algsete omaduste ja A-vitamiini kadumise. Märkus: kuumutustemperatuur steriliseerimise ajal on 120 kraadi.

Pastöriseerimisel, et toitaineid võimalikult palju säilitada, kuumutatakse piim vaid 80-90 kraadini. See mõjutab säilivusaega: piim rikneb kiiresti, kuna pastöriseerimise ajal eosed ei sure, vaid vastupidi, idanevad ja hakkavad kiiresti paljunema, mistõttu toode rikneb.
Sporulatsioon sporogenees, eoste moodustumise protsess (vt eosed). Taimeorganismides - prokarüootides (vt Prokarüootid), kelle rakkudel puuduvad tüüpilised tuumad, võivad eosed tekkida: tervest rakust, mis on kogunud toitaineid ja paksendanud kesta (paljude sinivetikate eksospoorid); Protoplast a jagamisel suureks hulgaks eosteks (mõnede sinivetikate endospoorid, riis. 1,
1); rakumembraani sees oleva protoplasti tihendamise ja kokkusurumise ning selle peale uue mitmekihilise membraani moodustumise tulemusena (bakterites); kui seeneniidistiku eriosad lagunevad segmentideks (aktinomütseedides, riis. 1
, 2). Taimedes - eukarüootides (vt. Eukarüootid), millel on tüüpilised tuumad, on 3 põhitüüpi eosed (oo-, mito- ja meiospoorid) ja mis asuvad arengutsüklites erinevates kohtades, võib esineda vastavalt 3 eoste varianti: oosporogenees. , mitosporogenees ja meosporogenees . Tavaliselt viitab S. meiospooride tekkele (meiosporogenees). Oosporogenees on seotud viljastamisprotsessiga ja seega ka tuumafaaside muutumisega arengutsüklites; lõpeb oospooride (paljudes rohevetikates ja oomütseetides), auksospooride (diatomites) ja sügospooride (sigomütseetides) moodustumisega, mis on ühe- või mitmetuumalised sigootid. Mitosporogenees põhjustab mitospooride teket, mida moodustuvad mitmel või suurel hulgal haploidide mitootilise jagunemise (vt Mitoos) tulemusena [näiteks mitmete vetikate zoospoorid ( riis. 1
, 3) ja seened], harvemini diploidsed (näiteks enamiku floriidide karpospoorid) rakud või jagunemata - edogooniumi monospoorid ( riis. 1
, 4), Bangiaceae, Nemalionaceae; ei too kaasa tuumafaaside muutust. Seda esineb üherakulistes mitosporangiumides (näiteks ulotriksi zoosporangiumides, edogooniumi monosporangiumides, floridia tsüstokarpides) ja ainuraksed vetikad ise muutuvad sporangiumideks ( riis. 1
, 5). Mitosporogeneesi võib täheldada dikaroone sisaldavatest rakkudest koosneva seeneniidistiku lagunemise ajal, näiteks täkk- ja roosteseentes. Meiosporogenees on seotud diplofaasi muutumisega (vt Diplofaas) nii madalamate kui ka kõrgemate taimede arengutsüklites haplofaasi teel (vt Haplofaas). Madalamates taimedes tekivad meioosi tagajärjel või vahetult pärast seda meioosi käigus tekkinud mitootiliselt jagunenud haploidrakkudest meiospoorid. Haploidse arengutsükliga vetikatel ja seentel toimub sulandumine sügoodi (oospooride) idanemisel, mille diploidne tuum moodustab meiootiliselt jagunedes 4 haploidset tuuma; sel juhul tekib 4 meiospoori (näiteks klamüdomonase zoospoorid, riis. 1
, 6, Ulotrixi aplanospoorid) või 3 neljast haploidsest tuumast surevad ja moodustub ainult 1 meiospoor (näiteks Spirogyras, riis. 1
, 7), või meioosile järgneb 1-3 mitootilist jagunemist ja moodustub 8-32 spoori (näiteks Bangiaceae). Isomorfse ja heteromorfse arengutsükliga vetikates toimub meiosporogenees üherakulistes meiosporangiumides ja seda iseloomustab kas 4 meiospoori moodustumine (näiteks pruunvetikate tetraspoorid ja enamik floriididest, riis. 1
, 8) või 16-128 meiospoori (näiteks pruunvetika zoospoorid, riis. 1
, 9) meioosile järgnenud 2-5 mitootilise jagunemise tõttu. Marsupiaalsete seente sporangiumides (kotid ehk asci) jagunevad mitootiliselt 4 meioosi tagajärjel tekkivat haploidset tuuma ja moodustub 8 endogeenset meiospoori (askospoori). Basiidiomütseedide basiidiatesse (eosed kandvatesse elunditesse) tekivad pärast meioosi 4 haploidset tuuma, mis liiguvad basiidiumi pinnale spetsiaalsetesse väljakasvudesse; tulevikus need haploidsete tuumadega väljakasvud, s.o. basidiospoorid eraldatakse basiidiatest ( riis. 1
, 10). Kõrgemad taimed moodustavad ainult meiospoore, meiosporogenees toimub mitmerakulistes sporangiumides. Tavaliselt tekib archesporiumi (vt Archesporium) diploidsete rakkude mitootiliste jagunemiste tulemusena nn. sporotsüüdid (meiootiliselt jagunevad rakud), moodustades 4 eost (eoste tetrad). Homospoorsed pteridofüüdid toodavad morfoloogiliselt ja füsioloogiliselt identseid eoseid ( riis. 2
, 1), millest arenevad biseksuaalsed võrsed. Heterospoorsetes sõnajalgades ja seemnetaimedes toimub mikro- ja megasporogenees ning meiosporogenees, st tekivad kahte tüüpi eosed. Mikrosporogenees toimub mikrosporangiates ja lõpeb suure hulga mikrospooride moodustumisega ( riis. 2
, 2), seejärel tärkavad isased võrsed; megasporogenees - megasporangiates, kus küpsevad väiksemal arvul - sageli isegi 4 või 1 - megaspoorid ( riis. 2
, 3), võrsudes emasteks võrseteks. Arenevad sporotsüüdid ja eosed (enamikus kõrgemates taimedes) toituvad ainetest, mis on saadud tapetumi rakkudest (sporangiumi õõnsust seestpoolt vooderdav kiht). Paljudes taimedes moodustavad selle kihi rakud laiali levides periplasmoodiumi (degenereeruvate tuumadega protoplasmaatiline mass), millesse ilmuvad sporotsüüdid ja seejärel eosed. Mõnedes taimedes osalevad periplasmoodiumi moodustamises ka mõned sporotsüüdid. Mõnede katteseemnetaimede megasporangiumides (munarakkudes) moodustuvad meioosi tagajärjel 2 või 4 haploidse tuumaga rakud, mis vastavad 2 ( riis. 2
, 4) või 4 ( riis. 2
, 5) megaspoorid; Nendest rakkudest arenevad emased gametofüüdid – nn. bispoorsed ja tetraspoorsed embrüokotid. S. kohta algloomades vt Art. Vaidlused. Lit.: Meyer K.I., Taimede paljundamine, M., 1937; Kursanov L.I., Komarnitsky N.A., Madalamate taimede kurss, M., 1945; Mageshwari P., Angiospermide embrüoloogia, tlk. inglise keelest, M., 1954; Takhtadzhyan A.L., Kõrgemad taimed, 1. kd, M. - L., 1956; Poddubnaya-Arnoldi V.A., Üldembrüoloogia katteseemnetaimedest, M., 1964: Smith G. M., Cryptogamic botany, 2 ed., v. 1-2, N. Y. - L., 1955; Lehrbuch der Botanik für Hochschulen, 29 Aufl., Jena, 1967. A. N. Sladkov. Riis. 1. Sporulatsioon madalamatel taimedel. 1 - endospooride moodustumine ja vabanemine sinivetikas Dermocarpa; 2 - seeneniidistiku lagunemine segmentideks aktinomütseedis Nocardia; 3 - ulothrix: eoste (a) ja eoste (b) vabanemine; 4 - edogoonium (Oedogonium): zoospooride vabanemine; 5 – Chlamydomonas: neli eost neid tootva raku kestas; 6 - klamüdomonas (Chlamydomonas): sügoot (a) ja selle idanevus nelja eosega (b); 7 - spirogyra (Spirogyra): sügoot (a) ja selle idanemine - nelja haploidse tuuma moodustumine (b), kolme tuuma surm (c), ühetuumaline seemik (d); 8 - Callithamnion: tetrasporangium (a) ja tetraspooride väljapääs (b); 9 - pruunvetikas Chorda filum: diploidse tuumaga sporangium (a), nelja (b) ja kuueteistkümne (c) haploidse tuumaga, peaaegu küpsete eostega (d); 10 - basiidium koos dikarüooniga (a), diploidse tuumaga (b) ja nelja haploidse tuumaga (c) basidiomütseetides; d - haploidse tuuma üleminek basidiospooriks. Riis. 2. Sporulatsioon kõrgemates taimedes. 1 - sporangiumi areng homospoorsel leptosporangiaatsel sõnajalgal; 2 - mikrosporangiumi areng Selaginellas; 3 - mikrosporangiumi areng asoolias (Azolla); 4 - megasporotsüüt (a) enne meioosi ja sellest tekkivad rakud pärast meioosi esimest (b) ja teist (c) jagunemist sibulas (Allium cepa); 5 - megasporotsüüt (a) enne meioosi ning kahetuumalised (b) ja neljakordsed (c, d) rakud, mis tekkisid pärast esimest ja teist meioosi jagunemist sarapuukull (Fritillaria persica). AC - archesporium, TP - tapetum, PP - periplasmodium, SC - sporotsüüdid, JS - eoste tuumad, SP - eosed (isospoorid), MS - megaspoor, TM - mikrospooride tetrad.
Suur Nõukogude entsüklopeedia. - M.: Nõukogude entsüklopeedia. 1969-1978 .
Sünonüümid:Vaadake, mis on "eoste moodustumine" teistes sõnaraamatutes:
Sporulatsioon... Õigekirjasõnastik-teatmik
Eosed on puhkavate, st grampositiivse rakuseina struktuuriga bakterite omapärane vorm. Eosed tekivad bakterite eksisteerimiseks ebasoodsates tingimustes (kuivamine, toitainete defitsiit jne. Bakteriraku sees tekib üks eos (endospoor) Eoste teke aitab kaasa liigi säilimisele ja ei ole paljunemisviis nagu seened.Perekonna Bacillus spoore moodustavate bakterite eosed ei ületa raku läbimõõtu Baktereid, mille eoste suurus ületab raku läbimõõdu, nimetatakse klostriidideks, näiteks perekonna Clostridium (lat. Clostridium) bakterid - spindli).
Spooride moodustumise protsess läbib mitmeid järjestikuseid etappe:
ettevalmistav Ainevahetus muutub, DNA replikatsioon lõpeb ja toimub kondenseerumine. Rakk sisaldab kahte või enamat nukleoidi, millest üks paikneb eoste tsoonis, ülejäänud on sporangiumi tsütoplasmas. Samal ajal sünteesitakse dipikoliinhapet;
prespore staadium. Vegetatiivse raku tsütoplasmaatilise membraani küljel toimub kahekordse membraani ehk vaheseinte sissekasv, mis eraldab nukleoidi tihendatud tsütoplasma alaga (sporogeenne tsoon). Selle tulemusena moodustub prospoor, mis on ümbritsetud kahe membraaniga;
kesta moodustumine. Esiteks moodustub prospoori membraanide vahele algeline peptidoglükaanikiht, seejärel ladestub selle kohale ajukoore paks peptidoglükaanikiht ja selle välismembraani ümber moodustub spoorimembraan;
eoste küpsemine. Kõikide eosstruktuuride moodustumine on lõpule viidud, see muutub kuumakindlaks, omandab iseloomuliku kuju ja hõivab rakus teatud positsiooni.
15. Komplekssed värvimismeetodid. Gramvärvimine, selle etapid.
Kandke preparaadile järjekindlalt teatud värvaineid, mis erinevad keemilise koostise ja värvuse poolest, peitsi, alkohole, happeid jne. See võimaldab tuvastada rakustruktuure ja eristada teatud tüüpi mikroorganisme teistest.
Grami plekk.
1. Kandke preparaadi fikseeritud määrdumisele emajuurviolett karboolalkoholi lahust läbi filterpaberi riba. 1-2 minuti pärast eemaldage see ja peske värvaine maha.
2. Kandke Lugoli lahust 1-2 minutiks
3. Värvige etüülalkoholiga 30-6 sekundit, kuni violetsed värvijoad lakkavad väljumast.
4. Loputage veega.
5. Viimistlege fuksia vesilahusega 1-2 minutit, loputage, kuivatage ja uurige mikroskoobiga. Gram+: tumelilla värv. Gramm -: punane
16. Ziehl-Neelseni värvimine ja selle etapid.
Ziehl-Neelseni värvimismeetod on mikroorganismide värvimismeetod happekindlate mükobakterite (tuberkuloosi, mükobakterioosi, leepra patogeenide), aktinomütseedide ja teiste happekindlate mikroorganismide tuvastamiseks. Mikroorganismide happekindlus on tingitud lipiidide, vahade ja hüdroksühapete olemasolust nende rakkudes. Sellised mikroorganismid on lahjendatud värvainete lahustega halvasti värvitud. Et hõlbustada värvaine tungimist mikroorganismide rakkudesse, kuumutatakse preparaadile kantud Zili karboolfuksiini põleti leegi kohal. Mineraalhapete ja alkoholi nõrgad lahused ei muuda värvilisi mikroorganisme.
Põleti leegile kinnitatud määrdumine määritakse 3–5 minutiks. Ziehli karboolfuksia või fuksiiniga värvitud paberi lahusega, kuumutatakse kuni auru ilmumiseni, kuid ilma värvi keemiseni.
Laske preparaadil jahtuda, eemaldage paber, nõrutage liigne värvaine maha ja peske preparaat veega.
Värvilist preparaati värvitustatakse 5% väävelhappe lahusega 3–5 s või 96° etüülalkoholiga, mis sisaldab 3 mahuprotsenti vesinikkloriidhapet, sukeldades mitu korda klaasi määrdiga soolhappealkoholiga klaasi.
Pärast värvi eemaldamist ülejäänud hape kurnatakse ja preparaat pestakse põhjalikult veega.
Täiendav värvimine toimub 3–5 minuti jooksul Loeffleri metüleensinisega.
Värvilist preparaati pestakse veega, kuivatatakse ja uuritakse mikroskoopiliselt.
Preparaatide värvimisel värvitakse happekindlad bakterid fuksiiniga rubiinpunast värvi ja hape ei muuda nende värvi.
Mittehappekindlad bakterid, samuti koeelemendid ja leukotsüüdid muutuvad happe mõjul värviliseks ja omandavad täiendava värvaine värvuse.
Sektsiooni on väga lihtne kasutada. Lihtsalt sisestage soovitud sõna vastavale väljale ja me anname teile selle tähenduste loendi. Tahaksin märkida, et meie sait pakub andmeid erinevatest allikatest - entsüklopeedilistest, selgitavatest, sõnamoodustussõnastikest. Siin näete ka näiteid sisestatud sõna kasutamisest.
Sõna sporulatsioon tähendus
sporulatsioon ristsõnasõnaraamatus
Sporulatsioon
sporogenees, eoste moodustumise protsess. Taimeorganismides - prokarüootides, kelle rakkudel puuduvad tüüpilised tuumad, võivad eosed tekkida: tervest rakust, mis on kogunud toitaineid ja paksendanud kesta (paljude sinivetikate eksospoorid); protoplasti jagamisel suureks hulgaks eosteks (mõnede sinivetikate endospoorid, riis. 1,
; rakumembraani sees oleva protoplasti tihendamise ja kokkusurumise ning selle peale uue mitmekihilise membraani moodustumise tulemusena (bakterites); kui seeneniidistiku eriosad lagunevad segmentideks (aktinomütseedides, riis. 1,
Taimedel - eukarüootidel, millel on tüüpilised tuumad, on 3 peamist tüüpi eoseid (oo-, mito- ja meiospoorid) ja nad hõivavad arengutsüklites erinevaid kohti, eoste variante võib olla vastavalt 3: oosporogenees, mitosporogenees ja meiosporogenees. . Tavaliselt viitab S. meiospooride tekkele (meiosporogenees). Oosporogenees on seotud viljastamisprotsessiga ja seega ka tuumafaaside muutumisega arengutsüklites; lõpeb oospooride (paljudes rohevetikates ja oomütseetides), auksospooride (diatomites) ja sügospooride (sigomütseetides) moodustumisega, mis on ühe- või mitmetuumalised sigootid. Mitosporogenees põhjustab mitospooride teket, mida moodustuvad mitmel või suurel hulgal haploidide mitootilise jagunemise (vt Mitoos) tulemusena [näiteks mitmete vetikate zoospoorid ( riis. 1,
ja seened], harvemini diploidsed (näiteks enamiku floriidide karpospoorid) või jagunemata ≈ edogooniumi monospoorid ( riis. 1,
Bangiaceae, nemalionaceae; ei too kaasa tuumafaaside muutust. Seda esineb üherakulistes mitosporangiumides (näiteks ulotriksi zoosporangiumides, edogooniumi monosporangiumides, floridia tsüstokarpides) ja ainuraksed vetikad ise muutuvad sporangiumideks ( riis. 1,
Mitosporogeneesi võib täheldada dikaroone sisaldavatest rakkudest koosneva seeneniidistiku lagunemise ajal, näiteks täkk- ja roosteseentes. Meiosporogenees on seotud diplofaasi muutumisega nii madalamate kui ka kõrgemate taimede arengutsüklites haplofaaside kaupa. Madalamates taimedes tekivad meioosi tagajärjel või vahetult pärast seda meioosi käigus tekkinud mitootiliselt jagunenud haploidrakkudest meiospoorid. Haploidse arengutsükliga vetikatel ja seentel toimub sulandumine sügoodi (oospooride) idanemisel, mille diploidne tuum moodustab meiootiliselt jagunedes 4 haploidset tuuma; sel juhul tekib 4 meiospoori (näiteks klamüdomonase zoospoorid, riis. 1, 6, Ulotrixi aplanospoorid) või 3 neljast haploidsest tuumast surevad ja moodustub ainult 1 meiospoor (näiteks Spirogyras, riis. 1, 7) või meioosile järgneb 1≈3 mitootilist jagunemist ja moodustub 8≈32 eost (näiteks Bangiaceae puhul). Isomorfse ja heteromorfse arengutsükliga vetikates toimub meiosporogenees üherakulistes meiosporangiumides ja seda iseloomustab kas 4 meiospoori moodustumine (näiteks pruunvetikate tetraspoorid ja enamik floriididest, riis. 1, 8) või 16≈128 meiospoori (näiteks pruunvetika zoospoorid, riis. 1, 9) 2≈5 mitootilise jagunemise tõttu pärast meioosi. Marsupiaalsete seente sporangiumides (kotid ehk asci) jagunevad mitootiliselt 4 meioosi tagajärjel tekkivat haploidset tuuma ja moodustub 8 endogeenset meiospoori (askospoori). Basiidiomütseedide basiidiatesse (eosed kandvatesse elunditesse) tekivad pärast meioosi 4 haploidset tuuma, mis liiguvad basiidiumi pinnale spetsiaalsetesse väljakasvudesse; tulevikus need haploidsete tuumadega väljakasvud, s.o. basidiospoorid eraldatakse basiidiatest ( riis. 1, 10). Kõrgemad taimed moodustavad ainult meiospoore, meiosporogenees toimub mitmerakulistes sporangiumides. Tavaliselt tekib diploidsete archespoorsete rakkude mitootiliste jagunemiste tulemusena nn. sporotsüüdid (meiootiliselt jagunevad rakud), moodustades 4 eost (eoste tetrad). Homospoorsed pteridofüüdid toodavad morfoloogiliselt ja füsioloogiliselt identseid eoseid ( riis. 2, 1), millest arenevad biseksuaalsed võrsed. Heterospoorsetes sõnajalgades ja seemnetaimedes toimub mikro- ja megasporogenees ning meiosporogenees, st tekivad kahte tüüpi eosed. Mikrosporogenees toimub mikrosporangiates ja lõpeb suure hulga mikrospooride moodustumisega ( riis. 2, 2), seejärel tärkavad isased võrsed; megasporogenees ≈ megasporangiates, kus väiksemal arvul ≈ sageli küpseb isegi 4 või 1 ≈ megaspoore ( riis. 2, 3), võrsudes emasteks võrseteks. Arenevad sporotsüüdid ja eosed (enamikus kõrgemates taimedes) toituvad ainetest, mis on saadud tapetumi rakkudest (sporangiumi õõnsust seestpoolt vooderdav kiht). Paljudes taimedes moodustavad selle kihi rakud laiali levides periplasmoodiumi (degenereeruvate tuumadega protoplasmaatiline mass), millesse ilmuvad sporotsüüdid ja seejärel eosed. Mõnedes taimedes osalevad periplasmoodiumi moodustamises ka mõned sporotsüüdid. Mõnede katteseemnetaimede megasporangiumides (munarakkudes) moodustuvad meioosi tagajärjel 2 või 4 haploidse tuumaga rakud, mis vastavad 2 ( riis. 2, 4) või 4 ( riis. 2, 5) megaspoorid; Nendest rakkudest arenevad emased gametofüüdid – nn. bispoorsed ja tetraspoorsed embrüokotid. S. kohta algloomades vt Art. Vaidlused.
Lit.: Meyer K.I., Taimede paljundamine, M., 1937; Kursanov L.I., Komarnitsky N.A., Madalamate taimede kurss, M., 1945; Mageshwari P., Angiospermide embrüoloogia, tlk. inglise keelest, M., 1954; Takhtadzhyan A.L., Kõrgemad taimed, 1. kd, M. ≈ L., 1956; Poddubnaya-Arnoldi V.A., Üldembrüoloogia katteseemnetaimedest, M., 1964: Smith G. M., Cryptogamic botany, 2 ed., v. 1≈2, N. Y. ≈ L., 1955; Lehrbuch der Botanik für Hochschulen, 29 Aufl., Jena, 1967.
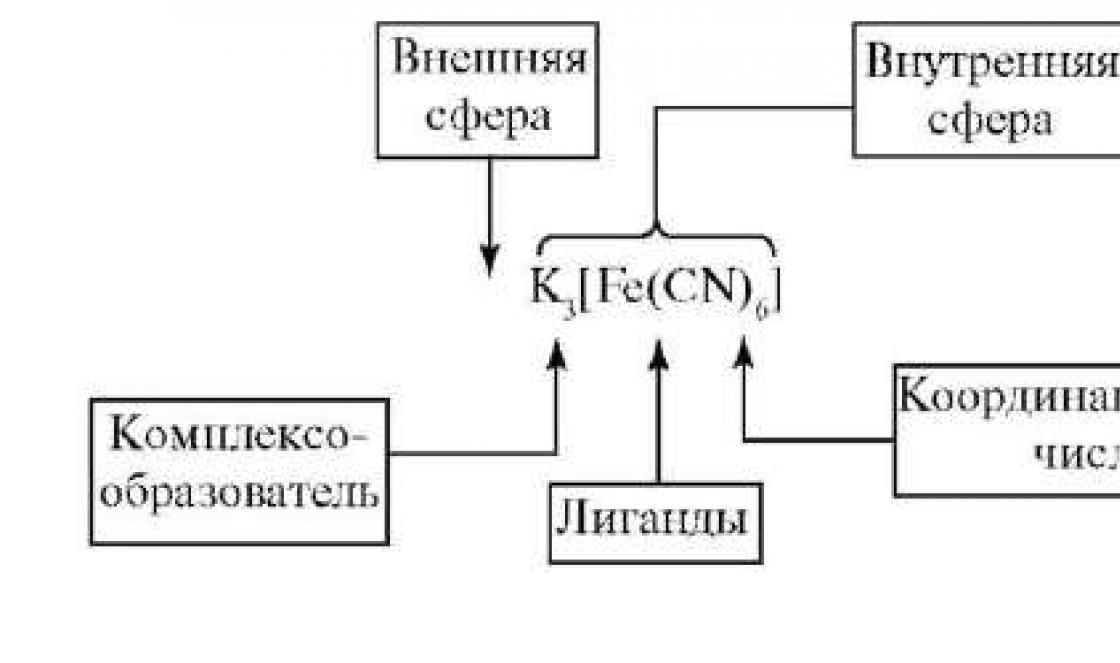
Vaidlused - puhkeolekus olevate grampositiivsete bakterite vorm. Eosed tekivad bakterite eksisteerimiseks ebasoodsates tingimustes (kuivamine, toitainete puudus jne). Sel juhul moodustub ühe bakteri sees üks spoor. Eoste moodustumine aitab kaasa liigi säilimisele ega ole paljunemisviis. Spoore moodustavaid pulgakujulisi aeroobseid baktereid, mille eoste suurus ei ületa raku läbimõõtu, nimetatakse batsillideks. Eoseid moodustavaid pulgakujulisi anaeroobseid baktereid, mille spooride suurus ületab bakteriraku, nimetatakse klostriidideks.
Eoste moodustumise skeem (G. Schlegeli järgi). A ja B – vaheseina moodustumine. C ja D – spoori protoplasti ümbritseb emaraku membraan. D – ajukoore ja eosmembraanide moodustumine. E – küpse spoori ehituse skeem: 1 – tsütoplasma nukleoidiga; 2 – CM eosed; 3 – eoste rakusein; 4 – ajukoor; 5 – eoste sisekest; 6 – eoste väliskest; 7 – eksospoorium.
Sporulatsiooni protsess(sporulatsioon) läbib mitmeid etappe. Esiteks, bakteriraku ühel poolusel nukleoid kondenseerub ja eraldub vaheseina moodustumise tõttu. Seejärel hakkab CPM moodustunud spoori protoplasti üle kasvama ja tekib volt, mis koosneb kahest CPM-i kihist, hiljem need ühinevad, mille tulemusena moodustunud eespoor on ümbritsetud kahekordse kestaga. Üksteise vastas olevate membraanide vahele moodustub embrüonaalne sein, ajukoor ning membraanidest väljaspool paiknevad välis- ja sisemembraanid.