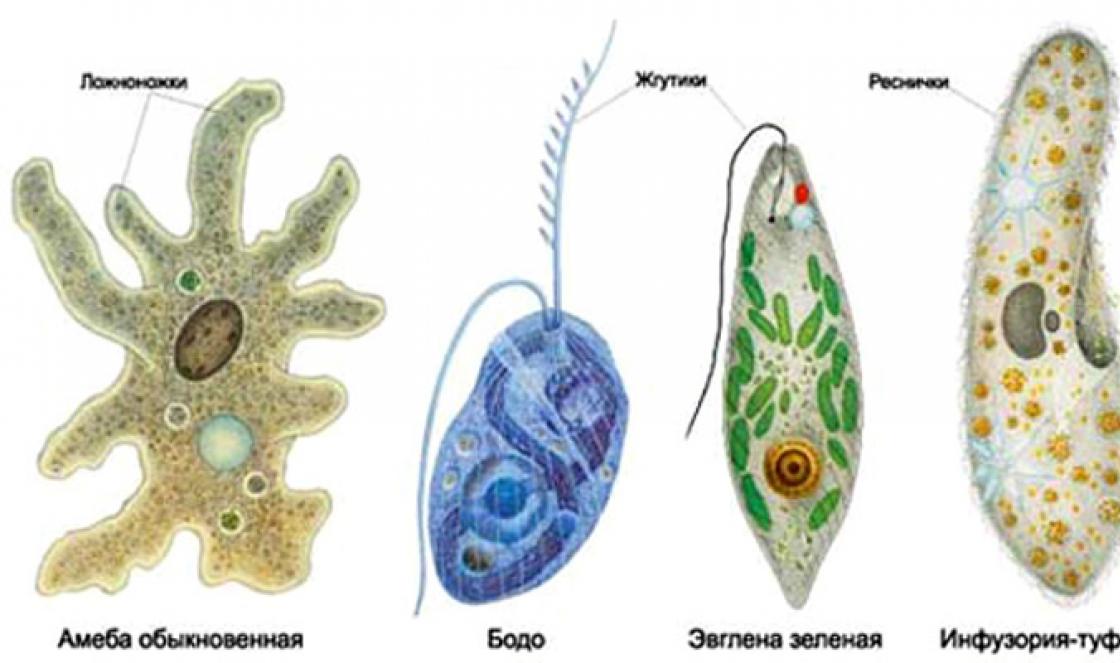
A mikroorganizmusok (mikrobák) 0,1 mm-nél kisebb méretű egysejtűek, amelyek szabad szemmel nem láthatók. Ide tartoznak a baktériumok, mikroalgák, néhány alsóbbrendű fonalas gomba, élesztő és protozoa (1. ábra). A mikrobiológia tanulmányozza őket.
Rizs. 1. Mikrobiológiai objektumok.
ábrán. 2. Az egysejtű protozoák néhány képviselője látható. Néha ennek a tudománynak a tárgyai közé tartoznak a Föld legprimitívebb szervezetei - olyan vírusok, amelyek nem rendelkeznek sejtszerkezettel, és nukleinsavak (genetikai anyag) és fehérje komplexei. Gyakrabban elkülönítik őket egy teljesen külön kutatási területre (Virológia), mivel a mikrobiológia inkább a mikroszkopikus egysejtű szervezetek tanulmányozására irányul.

Rizs. 2. Az egysejtű eukarióták (protozoák) egyes képviselői.
Az algák és gombák vizsgálatával foglalkozó algológia és mikológia tudományok külön tudományágak, amelyek átfedésben vannak a mikrobiológiával a mikroszkopikus élő objektumok vizsgálatában. A bakteriológia a mikrobiológia igazi ága. Ez a tudomány kizárólag a prokarióta mikroorganizmusok vizsgálatával foglalkozik (3. ábra).

Rizs. 3. Prokarióta sejt vázlata.
Az eukariótáktól eltérően, amelyek magukban foglalják az összes többsejtű szervezetet, valamint a protozoonokat, mikroszkopikus algákat és gombákat, a prokariótáknak nincs kialakult magja, amely genetikai anyagot és valódi organellumokat (a sejt állandó speciális struktúráit) tartalmazna.
A prokarióták közé tartoznak a valódi baktériumok és archaeák, amelyeket a modern besorolás szerint doméneknek (szuperkirályságoknak) neveznek Archaea és Eubacteria (4. ábra).

Rizs. 4. A modern biológiai osztályozás területei.
A baktériumok szerkezeti jellemzői
A baktériumok fontos láncszemei a természetben előforduló anyagok körforgásában, lebontják a növényi és állati maradványokat, megtisztítják a szerves anyagokkal szennyezett víztesteket, módosítják a szervetlen vegyületeket. Nélkülük nem létezhetne élet a földön. Ezek a mikroorganizmusok mindenhol elterjedtek, talajban, vízben, levegőben, állati és növényi szervezetekben.
A baktériumok a következő morfológiai jellemzőkben különböznek egymástól:
- Sejtforma (kerek, rúd alakú, fonalas, tekercses, spirális, valamint különféle átmeneti lehetőségek és csillag alakú konfiguráció).
- Mozgást segítő eszközök jelenléte (mozgásképtelen, lobogó, nyálkakiválasztás miatt).
- A sejtek egymás közötti artikulációja (izolált, párok, granulátumok, elágazó formák formájában összekapcsolva).
A gömbölyű baktériumok (coccusok) által alkotott struktúrák között vannak olyan sejtek, amelyek az osztódás után párban állnak, majd egyes képződményekre bomlanak (mikrokokkuszok), vagy folyamatosan együtt maradnak (diplococcusok). A négy sejtből álló négyzetes szerkezetet a tetracoccusok, a láncot a streptococcusok, a 8-64 egységből álló granulátumot a sarcina, a klasztereket a staphylococcusok alkotják.
A pálcika alakú baktériumok változatos alakban jelennek meg a sejt hossza (0,1-15 µm) és vastagsága (0,1-2 µm) nagy eltérése miatt. Ez utóbbi alakja attól is függ, hogy a baktériumok képesek-e spórákat képezni - olyan vastag héjú szerkezeteket, amelyek lehetővé teszik a mikroorganizmusok számára, hogy túléljék a kedvezőtlen körülményeket. Az ezzel a képességgel rendelkező sejteket bacilusoknak, az ilyen tulajdonságokkal nem rendelkező sejteket pedig egyszerűen pálcika alakú baktériumoknak nevezzük.
A pálcika alakú baktériumok speciális módosulatai a fonalas (megnyúlt) formák, láncok és elágazó szerkezetek. Ez utóbbit az aktinomicéták képezik egy bizonyos fejlődési szakaszban. Az „ívelt” rudakat hullámos baktériumoknak nevezik, amelyek között vibriókat izolálnak; két ívű spirilla (15-20 µm); hullámos vonalakra emlékeztető spirocheták. Sejthosszuk 1-3, 15-20 és 20-30 µm. ábrán. Az 5. és 6. ábra a baktériumok főbb morfológiai formáit, valamint a sejtben a spórák elrendezésének típusait mutatja be.

Rizs. 5. A baktériumok alapformái.

Rizs. 6. Baktériumok a sejtben a spórák elhelyezkedésének típusa szerint. 1, 4 – középen; 2, 3, 5 – véghely; 6 – oldalról.
A baktériumok fő sejtszerkezetei: nukleoid (genetikai anyag), fehérjeszintézisre szánt riboszómák, citoplazmatikus membrán (a sejtmembrán egy része), amely sok képviselőben felülről is védett, kapszula és nyálkahártya (7. ábra).

Rizs. 7. Egy baktériumsejt vázlata.
A baktériumok osztályozása szerint több mint 20 típus létezik. Például rendkívül termofil (magas hőmérsékletet kedvelők) Aquificae, anaerob pálca alakú baktériumok Bacteroidetes. A legdominánsabb törzs azonban, amely változatos képviselőket tartalmaz, az Actinobacteria. Ide tartoznak a bifidobaktériumok, a laktobacillusok és az aktinomicéták. Ez utóbbi egyedisége abban rejlik, hogy egy bizonyos fejlődési szakaszban micéliumot képez.
A köznyelvben ezt micéliumnak hívják. Valójában az aktinomyceták elágazó sejtjei gombahifákra hasonlítanak. Ennek ellenére az aktinomicétákat a baktériumok közé sorolják, mivel prokarióták. Természetesen sejtjeik szerkezetükben kevésbé hasonlítanak a gombákhoz.
Az Actinomycetes (8. ábra) lassan növekvő baktériumok, ezért nem képesek versenyezni a könnyen hozzáférhető szubsztrátumokért. Képesek lebontani olyan anyagokat, amelyeket más mikroorganizmusok nem tudnak szénforrásként felhasználni, különösen a kőolaj-szénhidrogéneket. Ezért az aktinomicétákat intenzíven tanulmányozzák a biotechnológia területén.
Egyes képviselők az olajmezők területére koncentrálnak, és speciális bakteriális szűrőt hoznak létre, amely megakadályozza a szénhidrogének behatolását a légkörbe. Az aktinomyceták gyakorlatilag értékes vegyületek aktív termelői: vitaminok, zsírsavak, antibiotikumok.

Rizs. 8. Reprezentatív aktinomyceta Nocardia.
Gombák a mikrobiológiában
A mikrobiológia tárgya csak az alsóbbrendű penészgombák (különösen a rhizopus, mucor). Mint minden gomba, nem képes önmagukban anyagokat szintetizálni, és tápközeget igényel. Ennek a királyságnak az alsóbb képviselőinek micéliuma primitív, nincs válaszfalakkal osztva. A mikrobiológiai kutatásban egy speciális rést foglal el az élesztő (9. ábra), amelyet a micélium hiánya jellemez.

Rizs. 9. Élesztőtenyészetek telepformái táptalajon.
Jelenleg sok ismeretet gyűjtöttek össze jótékony tulajdonságaikról. Az élesztőt azonban továbbra is tanulmányozzák gyakorlatilag értékes szerves vegyületek szintézisére való képessége miatt, és aktívan használják modellszervezetként genetikai kísérletekben. Az ókor óta az élesztőt az erjesztési folyamatokban használták. Az anyagcsere a különböző képviselők között eltérő. Ezért egyes élesztők alkalmasabbak egy adott folyamatra, mint mások.
Például a Saccharomyces beticust, amely jobban ellenáll a magas alkoholkoncentrációnak, erős borok készítésére használják (akár 24%). Míg a S. cerevisiae élesztő kisebb koncentrációjú etanolt képes előállítani. Felhasználási területük szerint az élesztőket takarmányozásra, pékekre, sörfőzdékre, szeszes italokra és borokra osztják.
Patogén mikroorganizmusok
Betegséget okozó vagy kórokozó mikroorganizmusok mindenhol megtalálhatók. A jól ismert vírusok: influenza, hepatitis, kanyaró, HIV stb. mellett veszélyes mikroorganizmusok a rickettsia, valamint a streptococcusok és a staphylococcusok, amelyek vérmérgezést okoznak. A pálcika alakú baktériumok között számos kórokozó található. Például diftéria, tuberkulózis, tífusz (10. ábra). Az emberre veszélyes mikroorganizmusok számos képviselője megtalálható a protozoonok között, különösen a maláriás plazmódium, a toxoplazma, a leishmania, a lamblia, a trichomonas és a patogén amőbák.

Rizs. 10. Fénykép a lépfenét okozó Bacillus anthracis baktériumról.
Sok aktinomyceta nem veszélyes emberre és állatra. A tuberkulózist és leprát okozó mikobaktériumok között azonban számos patogén képviselő található. Egyes aktinomicéták olyan betegséget indítanak el, mint például az aktinomikózis, amelyet granulomák képződése és néha a testhőmérséklet emelkedése kísér. A penészgombák bizonyos típusai képesek az emberre mérgező anyagokat - mikotoxinokat - termelni. Például az Aspergillus nemzetség néhány képviselője, a Fusarium. A patogén gombák a mikózisoknak nevezett betegségek csoportját okozzák. Így a candidiasist vagy egyszerűen fogalmazva a rigót élesztőszerű gombák okozzák (11. ábra). Mindig jelen vannak az emberi szervezetben, de csak akkor aktiválódnak, ha az immunrendszer legyengül.

Rizs. 11. A Candida gomba a rigó kórokozója.
A gombák különféle bőrelváltozásokat okozhatnak, különösen mindenféle zuzmót, kivéve a herpes zoster-t, amelyet vírus okoz. A Malassezia élesztő, az emberi bőr állandó lakója, csökkentheti az immunrendszer aktivitását. Ne rohanjon azonnal kezet mosni. Az egészséges élesztők és opportunista baktériumok fontos funkciót töltenek be, megakadályozva a patogén mikroorganizmusok kifejlődését.
A vírusok mint a mikrobiológia tárgya
A vírusok a legprimitívebb élőlények a Földön. Szabad állapotban nem mennek végbe bennük anyagcsere-folyamatok. A vírusok csak akkor kezdenek el szaporodni, amikor belépnek a gazdasejtbe. Minden élő szervezetben a genetikai anyag hordozója a dezoxiribonukleinsav (DNS). Csak a vírusok között vannak olyan genetikai szekvenciával rendelkező képviselők, mint a ribonukleinsav (RNS).
A vírusokat gyakran nem az igazán élő szervezetek közé sorolják.
A vírusok morfológiája igen változatos (12. ábra). Átmérőjük jellemzően 20-300 nm.

Rizs. 12. A vírusrészecskék sokfélesége.
Egyes képviselők 1-1,5 mikron hosszúságot érnek el. A vírus szerkezete abból áll, hogy a genetikai anyagot egy speciális fehérjevázzal (kapszid) veszi körül, amelyet sokféle forma (spirál, ikozaéder, gömb) jellemez. Egyes vírusok tetején a gazdasejt membránjából (superkapszid) képződött burok is található. Például (13. ábra) az úgynevezett (AIDS) betegség kórokozójaként ismert. Genetikai anyagként RNS-t tartalmaz, és az immunrendszer bizonyos típusú sejtjére (segítő T-limfocitákra) hat.

Rizs. 13. A humán immundeficiencia vírus szerkezete.
A baktériumok táplálkozása.
Táplálás.
Passzív diffúzió
Könnyített diffúzió
Aktiv szállitás
Az első esetben a tápanyagmolekula komplexet képez a periplazmatikus tér fehérjével, amely kölcsönhatásba lép a citoplazmatikus membrán egy specifikus permeázával. A citoplazma membránon keresztüli energiafüggő behatolás után a „szubsztrát - periplazmatikus fehérje - permeáz” komplex disszociál a szubsztrát molekula felszabadulásával.
Az aktív transzport során a szállított anyag kémiai módosításával az eseménylánc a következőket tartalmazza: (1) a membránenzim-2 foszforilációja a citoplazmából foszfoenolpiruváttal; (2) szubsztrát molekulák foszforilált enzim-2 általi kötődése a citoplazma membrán felületéhez; (3) a szubsztrát molekula energiafüggő transzportja a citoplazmába; (4) foszfátcsoport átvitele szubsztrát molekulára; (5) a „szubsztrát-enzim” komplex disszociációja a citoplazmában. A foszforiláció következtében a szubsztrát molekulák felhalmozódnak a sejtek citoplazmájában, és nem képesek elhagyni azokat.
A baktériumok osztályozása a táplálkozás típusa szerint.
A tápanyagfelvétel módja szerint anyagokra osztják a baktériumokat holofitákÉs holozoánok. Holofitikus baktériumok (görögből holos- teljes és phyticos– növényekkel kapcsolatos) nem képesek a szubsztrátokat lebontó enzimeket a környezetbe juttatni, ennek következtében a tápanyagokat kizárólag oldott, molekuláris formában fogyasztják. Holozoán baktériumok (görögből holos- teljes és zoikosz- az állatokkal kapcsolatos), éppen ellenkezőleg, exoenzim-komplexummal rendelkeznek, amelyek külső táplálkozást biztosítanak - a szubsztrátok lebontását a baktériumsejten kívüli molekulákká. Ezt követően a tápanyagmolekulák bejutnak a holozoán baktériumokba.
Szénforrás szerint között baktériumokat izolálnak autotrófokÉs heterotrófok. Autotrófok (görögből autók- magam, trófeát– élelmiszer) szén-dioxidot (CO 2) használnak szénforrásként, amelyből minden széntartalmú anyag szintetizálódik. Mert heterotrófok (a görög geteros szóból - egy másik és trófeát– élelmiszer) a szénforrás különböző szerves anyagok molekuláris formában (szénhidrátok, többértékű alkoholok, aminosavak, zsírsavak). A heterotrófia legnagyobb foka a prokariótákban rejlik, amelyek csak más élő sejtekben (például rickettsia és chlamydia) belül élhetnek.
Nitrogénforrással A prokarióták 3 csoportra oszthatók: 1) nitrogénmegkötő baktériumok (molekuláris nitrogént asszimilálnak a légköri levegőből); 2) olyan baktériumok, amelyek ammóniumsókból, nitritekből vagy nitrátokból szervetlen nitrogént fogyasztanak; 3) baktériumok, amelyek asszimilálják a szerves vegyületekben (aminosavak, purinok, pirimidinek stb.) található nitrogént.
Energiaforrás szerint a baktériumokat osztják fototrófokÉs kemotrófok. Fototróf baktériumok , mint a növények, képesek a napenergiát hasznosítani. A fototróf prokarióták emberben nem okoznak betegségeket. Kemotróf baktériumok redox reakciókból energiát nyernek.
Az elektrondonorok természeténél fogva litotrófok(görögből litos- kő) és organotrófok. U litotrófok (kemolitotrófok ) szervetlen anyagok (H 2, H 2 S, NH 3, kén, CO, Fe 2+ stb.) elektrondonorként működnek. Elektron donorok organotrófok (kemoorganotrófok ) szerves vegyületek – szénhidrátok, aminosavak stb.
A legtöbb emberre patogén baktérium kemoorganotróf (kemoheterotróf) típusú táplálkozással rendelkezik; A kemolitotróf (kemoautotróf) típus kevésbé gyakori.
Szerves vegyületek szintézisének képessége szerint a kemotróf baktériumokat osztják prototrófok, auxotrófokÉs hypotrophiák. Prototróf baktériumok szintetizálja az összes szükséges szerves anyagot glükózból és ammóniumsókból. A baktériumokat hívják auxotrófok , ha nem képesek szerves anyagot szintetizálni a meghatározott vegyületekből. Az anyagcsere-aktivitás extrém mértékű elvesztését alultápláltságnak nevezik. Hipotróf baktériumok létfontosságú funkcióikat a gazdaszervezet szerkezeteinek vagy metabolitjainak átszervezésével biztosítják.
A szén és nitrogén mellett a baktériumoknak kénre, foszforra és fémionokra van szükségük a teljes élethez. A kénforrások aminosavak (cisztein, metionin), vitaminok, kofaktorok (biotin, liponsav stb.), szulfátok. A foszfor forrásai a nukleinsavak, foszfolipidek és foszfátok. Megfelelően magas koncentrációban a baktériumoknak magnéziumra, káliumra, kalciumra és vasra van szükségük; sokkal kisebbekben - cink, mangán, nátrium, molibdén, réz, nikkel, kobalt.
Növekedési tényezők- ezek olyan anyagok, amelyeket a baktériumok nem tudnak maguktól szintetizálni, de nagy szükségük van rájuk. A növekedési faktorok közé tartoznak az aminosavak, nitrogéntartalmú bázisok, vitaminok, zsírsavak, vasporfirinek és más vegyületek. A baktériumok életéhez szükséges optimális feltételek megteremtése érdekében növekedési faktorokat kell hozzáadni a tápközeghez.
Anyagcsere, energiaátalakítás
A) Konstruktív anyagcsere.
A bakteriális táplálkozás kötelező szakasza a tápanyagok asszimilációja, azaz megváltoztatott vagy módosított formában történő beépítése a sejtkomponensek és struktúrák reprodukálására szolgáló szintetikus reakciókba.
Fehérje anyagcsere baktériumokban 3 fázisban fordulhat elő: elsődleges fehérjebontás, másodlagos lebontás és fehérjeszintézis. A fehérjemolekulák elsődleges lebontását peptonokká exoenzimek - a baktériumok által a környezetbe szekretált exoproteázok végzik. A másodlagos lebomlás endoenzimek (endoproteázok) hatására megy végbe, amelyekkel minden baktérium rendelkezik. Ez a folyamat a baktériumsejt belsejében megy végbe, és a peptidek aminosavakká történő lebontásából áll. Ez utóbbiak változatlanul felhasználhatók, vagy kémiai átalakulásoknak vethetők alá (dezaminálás, dekarboxilezés stb.), melynek eredményeként ammónia, indol, kénhidrogén, ketosavak, alkohol, szén-dioxid stb diagnosztikai értéke a bakteriológiában.
A fehérjék lebontási reakcióival együtt szintézisük reakciói is bekövetkeznek. Egyes baktériumok külső táplálkozás eredményeként kapott kész aminosavakból fehérjéket képeznek, míg más baktériumok önállóan szintetizálnak aminosavakat egyszerű, nitrogént és szenet tartalmazó vegyületekből. Az aminosavak szintézise aminálási, transzaminálási, amidálási és karboxilezési reakciókban valósítható meg. A legtöbb prokarióta képes szintetizálni a sejtfehérjéket alkotó összes aminosavat. Az aminosavak bioszintézisének sajátossága a közös bioszintézis utak alkalmazása: trikarbonsavciklus, glikolízis, oxidatív pentóz-foszfát út stb. Az aminosavak szintézisének fő kiindulási vegyületei a piruvát és a fumarát.
Szénhidrát anyagcsere különbözik az autotrófok és a heterotrófok között (1. séma). Az autotróf baktériumok szén-dioxidból szintetizálják az összes szükséges szénhidrátot. A heterotróf baktériumok szénhidrátképzésének nyersanyagai a következők lehetnek: (1) egy-, két- és három szénatomos vegyületek; és (2) poliszacharidok (keményítő, glikogén, cellulóz). Ez utóbbi lebontására számos heterotróf baktérium rendelkezik exoenzimekkel (amiláz, pektináz stb.), amelyek a poliszacharidokat hidrolizálva glükózt, maltózt, fruktózt stb.
Az autotróf baktériumokban a Calvin-ciklusban szén-dioxidból ribulóz-foszfát-foszfor-glicerinsav képződik, amely az ellenkező irányú glikolitikus reakciókban vesz részt. A fordított szintézis végterméke a glükóz.
A heterotróf baktériumok egy-, két- és háromszénatomos vegyületekből glükózt képeznek, bevonva őket a fordított glikolízis reakcióba is. Tekintettel arra, hogy a glikolízis egyes reakciói visszafordíthatatlanok, a heterotrófok speciális enzimreakciókat fejlesztettek ki, amelyek lehetővé teszik számukra, hogy megkerüljék a katabolikus útvonal irreverzibilis reakcióit.
Amikor a poliszacharidokat a heterotróf baktériumok lebontják, a keletkező diszacharidok bejutnak a sejtekbe, és maltóz, szacharóz és laktóz hatására hidrolízisen mennek keresztül, majd monoszacharidokká bomlanak, amelyeket azután fermentálnak, vagy beépítenek a cukor interkonverziós reakcióiba.
Lipid anyagcsere. Mind az exogén lipidek, mind az intersticiális metabolizmus amfibolitjai kiindulási anyagként szolgálhatnak a baktériumok lipidképződéséhez. Az exogén lipidek bakteriális lipázok és más lipolitikus enzimek hatásának vannak kitéve. Sokféle baktérium metabolizálja a glicerint, amely műanyag és energiaforrásként szolgál. A lipidszintézis endogén forrásai az acetil-koenzim A, a propionil-ACP, a malonil-ACP (ACP - acetiltranszfer protein), a foszfodioxi-aceton stb.
A páros szénatomszámú zsírsavak szintézisének kiindulási szubsztrátja az acetil-koenzim A, a páratlan szénatomszámú zsírsavak esetében - propionil-ACP és malonil-ACP. A kettős kötések kialakulása egy savmolekulában aerob prokariótákban a molekuláris oxigén és a deszaturáz enzim részvételével történik. Az anaerob prokariótákban a kettős kötések a szintézis korai szakaszában képződnek egy dehidratációs reakció eredményeként. A foszfolipidek szintézisének kiindulási szubsztrátja a foszfodioxi-aceton (a glikolitikus út köztiterméke), amelynek redukciója 3-foszforglicerin képződéséhez vezet. Utóbbihoz ezután 2 zsírsav-maradék kapcsolódik ACP-vel komplex formájában. A reakció terméke foszfatidsav, amelynek CTP-vel történő aktiválása, majd szerin, inozit, glicerin vagy más vegyület foszfátcsoporthoz való hozzáadása a megfelelő foszfolipidek szintéziséhez vezet.
A zsírsavakban auxotróf és hipotróf mikroorganizmusok (például mikoplazmák) ezeket kész formában nyerik ki gazdasejtekből vagy tápközegből.
Mononukleotid csere. A purin és pirimidin mononukleotidok a DNS és az RNS alapvető alkotóelemei. Sok prokarióta képes a tápközegben lévő kész purin és pirimidin bázisok, ezek nukleozidjainak és nukleotidjainak felhasználására, illetve kis molekulatömegű anyagokból történő szintetizálására. A baktériumok olyan enzimekkel rendelkeznek, amelyek az exogén purin- és pirimidinszármazékok interkonverziójának következő szakaszait katalizálják: nitrogénbázis – nukleozid – nukleotid (mono- – di- – trifoszfát).
Purin és pirimidin mononukleotidok szintézise de novo független módon hajtják végre. A purin nukleotidok szintézise során egymást követő enzimatikus reakciók eredményeként inozinsav képződik, amelyből a puringyűrű kémiai módosításával adenil (AMP) és guanilsav (GMP) szintetizálódik. A pirimidin nukleotidok szintézise az orotidinsav képződésével kezdődik, amelynek dekarboxilezése során uridilsav (UMA) keletkezik. Ez utóbbiból UTP képződik, melynek acilezése CTP képződéséhez vezet.
A dezoxiribonukleotidok a megfelelő ribonukleotidok difoszfátok vagy trifoszfátok szintjén történő redukciója eredményeként jönnek létre. A DNS-specifikus nukleotid, a timidilsav szintézise a dezoxiuridilsav enzimatikus metilezésén keresztül megy végbe.
Ioncsere. Ásványi vegyületek - ionok, NH 3 +, K +, Mg 2+, Fe 2+, SO 4 2-, PO 4 3- és más baktériumok szabadon és más szerves anyagokkal társított állapotban is nyerhetők a környezetből anyagokat. A kationok és anionok a 3. §-ban leírtak szerint különféle módokon jutnak be a baktériumsejtbe. Az ionok baktériumsejtbe való behatolási sebességét a környezet pH-ja és maguknak a mikroorganizmusoknak a fiziológiai aktivitása befolyásolja.
B) A baktériumok légzése (energia-anyagcsere).
Minden életfolyamat energiafüggő, ezért az energiatermelés a prokarióta anyagcsere rendkívül fontos aspektusa. Anaerob és aerob légzéssel nyernek energiát.
Lehelet, vagy A biológiai oxidáció egy katabolikus elektrontranszfer folyamata donor anyagból akceptor anyagba, amelyet energia felhalmozódása kísér nagy energiájú vegyületekben . A légzés katabolikus reakciók során megy végbe, melynek eredményeként az összetett szerves anyagok lebomlása során energiát szabadítanak fel és egyszerű vegyületekké alakulnak. A makroerg anyagokban (ATP, GTP, UTP stb.) felhalmozódott energiát anabolikus reakciókban hasznosítják.
A légzés módja szerint a mikroorganizmusokat felosztják kötelező (szigorú) aerobok, kötelező anaerobokÉs fakultatív anaerobok.
Kötelező aerobok szabad oxigénre van szükségük. Az emberre patogén kemoorganotróf aerobokban az elektrondonorok szerves vegyületek (szénhidrátok, zsírok, fehérjék), az elektronakceptor pedig a molekuláris oxigén. A kemoorganotróf aerobokban ATP formájában történő energiatárolás az elektrondonorok oxidatív foszforilációja során történik. Az aerobok citokrómokkal (az elektrontranszferben részt vesznek), valamint enzimekkel (kataláz, szuperoxid-diszmutáz, peroxidáz) rendelkeznek, amelyek inaktiválják a légzés során keletkező toxikus oxigéngyököket. A szuperoxid-diszmutáz inaktiválja a legmérgezőbb metabolitot, a H2O2 szuperoxidgyökét. A kataláz enzim a H 2 O 2 -t H 2 O 2 és O 2 -dá alakítja.
Az aerobok egy speciális csoportja mikroaerofil baktériumok amelyek bár oxigént igényelnek az energia megszerzéséhez, a megnövekedett CO 2 tartalommal jobban szaporodnak, például a nemzetséghez tartozó baktériumok CampylobacterÉs Helicobacter.
Kötelező anaerobok nincs szükségük szabad oxigénre, éppen ellenkezőleg, az oxigén még kis mennyiségben is mérgező hatással van rájuk. Különféle szerves vegyületek (főleg szénhidrátok) elektrondonorként szolgálnak az emberre patogén kemoorganotróf anaerobokhoz. A kemoorganotróf anaerobokban az elektronakceptor szerves oxigéntartalmú vegyületek - savak vagy ketonok, vagyis az elektronakceptor egy szerves fragmenshez kötött oxigén. Ezekben a prokariótákban az energia tárolása a szubsztrát foszforilációján keresztül történik. A kötelező anaerobok általában nem rendelkeznek citokrómokkal és enzimekkel, amelyek inaktiválják az oxigéngyököket (kataláz- és szuperoxid-diszmutáz-negatívak).
Az emberre nem patogén kemolitotróf anaerobokban az elektronakceptor szervetlen oxigéntartalmú vegyületek - nitrátok, szulfátok, karbonátok.
Az anaerobok egy speciális csoportját alkotják az aerotoleráns baktériumok, amelyek képesek szaporodni légköri oxigén jelenlétében, de nem használják azt elektronakceptorként (például tejsavbaktériumok). Az aerotoleráns prokarióták kataláz- és szuperoxid-diszmutáz-pozitívak.
Fakultatív anaerobok oxigén és oxigénmentes környezetben egyaránt képesek létezni. Elektrondonoraik szerves anyagok; elektronakceptorok, környezeti feltételektől függően - molekuláris oxigén vagy szerves és szervetlen vegyületekben kötött oxigén. Az energiát fakultatív anaerobok oxidatív és szubsztrát foszforilációval egyaránt felhalmozhatják. Az aerobokhoz hasonlóan ez a baktériumcsoport is tartalmaz citokrómokat és antioxidáns enzimeket.
Az energiatermelés fő szubsztrátja a szénhidrátok, amelyek a különböző légzéstípusú kemoheterotróf prokariótákban acetil-koenzim A-vá (aktivált ecetsavvá) katabolizálódhatnak. A lipidek és a fehérjék energiaszubsztrátként működhetnek, mivel az acetil-koenzim A szintén metabolizmusuk egyik köztes terméke (2. ábra).
A szénhidrát-katabolizmus kemoorganotróf prokariótákban a következőket foglalja magában: (a) anaerob folyamatok – glikolízis, pentóz-foszfát-útvonal és ketodezoxi-foszfoglükonát-útvonal; b) aerob folyamat – trikarbonsavciklus (Krebs-ciklus). Az anaerob folyamatok minden prokarióta esetében előfordulnak, míg az aerob folyamat csak az obligát aerobokra és a fakultatív anaerobokra jellemző. Az anaerob utakon történő energiatermelés a szubsztrát foszforiláción, míg az aerob folyamat az oxidatív foszforiláción alapul.
Fogalmak meghatározása.
A sterilizálás, a fertőtlenítés és az antiszeptikumok a modern orvosi és különösen a sebészeti gyakorlat szerves részét képezik. Ezen módszerek alapelveinek megértése és gyakorlati alkalmazása azért szükséges, mert sok potenciálisan patogén mikroorganizmus hosszú ideig életképes maradhat a gazdaszervezeten kívül, nagy ellenállást mutat a fizikai és kémiai fertőtlenítőszerekkel szemben, és viszonylag könnyen átvihető egyik emberről a másikra.
Antiszeptikumok- patogén vagy opportunista mikroorganizmusok elpusztítása vagy szaporodásának megakadályozása kémiai módszerekkel. Ezt a kifejezést általában egy kémiai gyógyszer élő szövetre történő külső alkalmazására használják.
Fertőtlenítő- olyan anyag, amely gátolja a mikroorganizmusok növekedését vagy elpusztítja (a baktériumspórák befolyásolása nélkül). A kifejezés olyan anyagokra vonatkozik, amelyeket élő szöveteken helyi hatásra használnak.
Fertőtlenítés a szepszis hiányát jelenti, de általában ezt a kifejezést az élő szervezetek hiányának hangsúlyozására használják. Aszeptikus technikák minden olyan eljárást jelent, amelyet az élő szervezetek kiirtására és az általuk történő újrafertőződés megelőzésére terveztek. A modern sebészeti és mikrobiológiai technikák aszeptikus eljárásokon alapulnak.
Biocid- olyan anyag, amely elpusztítja az összes élő mikroorganizmust, mind a patogén, mind a nem patogén, beleértve a spórákat is.
Biostat- olyan szer, amely megakadályozza a mikroorganizmusok szaporodását, de nem feltétlenül pusztítja el azokat.
Fertőtlenítés- a mikroorganizmusok eltávolítása mennyiségi meghatározás nélkül. Ez a kifejezés relatív; A csírák végső eltávolítása sterilizálással vagy fertőtlenítéssel történhet.
Fertőtlenítés- olyan folyamat, amely csökkenti vagy teljesen elpusztítja az összes kórokozó mikroorganizmust, kivéve a spórákat.
Csíraölő- olyan anyag, amely elpusztítja a mikroorganizmusokat, különösen a patogéneket. A csíraölő nem pusztítja el a spórákat.
Higiénia- olyan módszer, amellyel a mikrobiális szennyeződést „biztonságos” szintre csökkentik. Ezt a módszert korábban élettelen objektumok „tisztítására” használták.
Sterilizáció- fizikai tényezők és (vagy) vegyszerek használata a mikrobiális élet minden formájának teljes megsemmisítésére vagy megsemmisítésére.
Sterilizáció.
A sterilizálás az összes mikroorganizmus és spóráik megsemmisítése vagy eltávolítása (szűréssel). A sterilizálás általában hővel történik. A mikrobiológiai laboratórium működésének egyik rutin eljárásaként a sterilizálás szükséges módszer annak biztosítására, hogy a tenyészetek, berendezések, üvegedények és táptalajok csak a szükséges mikroorganizmusok szaporodását támogassák, míg a többi mikrobát elpusztítsák. A sterilizálásnak a következő típusai vannak: égő lángban történő kalcinálás, forralás, áramló gőzzel történő akció, nyomás alatti gőz autoklávban, száraz hő, pasztőrözés, tindizálás, kémiai, hideg (mechanikai) sterilizálás.
A sterilizációs módszerek kiválasztása.
A sterilizálási módszerek kiválasztásakor a következő követelményeket kell figyelembe venni:
1. Hatás: baktériumölő, sporicid, tuberkulocid, gombaölő és vírusölő.
2. Az eljárás gyorsasága: a sterilizálást a lehető leggyorsabban el kell végezni.
3. Permeabilitás: a sterilizáló anyagoknak a csomagoláson keresztül és a műszer belső részeibe kell behatolniuk.
4. Kompatibilitás: A többször sterilizált anyagok szerkezete vagy funkciója nem változhat.
5. Nem mérgező: nem jelenthet veszélyt az emberi egészségre vagy a környezetre.
6. Szerves anyag stabilitása: A sterilizálás hatékonysága nem csökkenhet szerves anyag jelenlétében.
7. Alkalmazhatóság: használható nagy és kis mennyiségű sterilizált anyaghoz.
8. Időbeli szabályozás: A feldolgozási ciklust könnyen és pontosan ellenőrizni kell.
9. Ár: a felszerelés, a telepítés és az üzemeltetés ésszerű költsége.
 Fizikai sterilizátorok
Fizikai sterilizátorok
Nedves meleg, amely a gőzzel végzett autoklávozás során keletkezik, a klinikai mikrobiológiai laboratóriumokban használt fő sterilizálószer. Az autoklávokat táptalajok, hőálló anyagok sterilizálására és fertőző hulladék kezelésére használják. A gőzsterilizáló vagy autokláv egy szigetelt, túlnyomásos kamra, amely telített gőzt használ a magas hőmérséklet létrehozására (1. ábra). A levegő eltávolítása a kamrából tömegeltolással vagy vákuum létrehozásával történik. A leggyakrabban használt autoklávok a tömeges helyettesítés. Könnyebb gőz kerül a kamrába, hogy kiszorítsa a nehezebb levegőt. A rövid nyomású gőzkezelés elpusztíthatja a baktériumspórákat. A táptalajok és egyéb anyagok rutinszerű sterilizálásához az expozíciós idő 15 perc 121°C-on, a nyomás pedig 1,5 kg/1 négyzetcentiméter. Fertőző hulladék esetén az expozíciós idő 30-60 percre nő. A megfelelő idő és hőmérséklet mellett a sterilizálás során nagyon fontos a gőzzel való közvetlen érintkezés. Fertőző anyag feldolgozásakor biztosítani kell a gőz maximális behatolását a hulladékba. Az ilyen anyagot 132ºС hőmérsékleten kell feldolgozni. Nem szabad autoklávozni daganatellenes gyógyszereket, mérgező vegyszereket és radioizotópokat, amelyek nem bomlanak le, valamint instabil vegyszereket, amelyek a hő hatására elpárologhatnak és szétterjedhetnek a kamrában.
 Száraz hő sterilizálás olyan anyagokhoz használják, amelyeket nem lehet gőzzel sterilizálni a sérülés lehetősége vagy az anyag gőzáteresztő képessége miatt. A száraz hő kevésbé hatékony, mint a nedves hő, és hosszabb expozíciós időt és magasabb hőmérsékletet igényel. A száraz hősterilizálást általában száraz hőkemencében végzik (2. ábra). A száraz hővel történő sterilizálás mechanizmusa oxidatív folyamat. Példák a száraz hővel sterilizáló anyagokra: olajok, porok, éles eszközök és üvegedények. Száraz hőt vagy termikus inaktiválást-sterilizálást alkalmaznak alternatív módszerként a fertőző hulladékok kezelésére.
Száraz hő sterilizálás olyan anyagokhoz használják, amelyeket nem lehet gőzzel sterilizálni a sérülés lehetősége vagy az anyag gőzáteresztő képessége miatt. A száraz hő kevésbé hatékony, mint a nedves hő, és hosszabb expozíciós időt és magasabb hőmérsékletet igényel. A száraz hősterilizálást általában száraz hőkemencében végzik (2. ábra). A száraz hővel történő sterilizálás mechanizmusa oxidatív folyamat. Példák a száraz hővel sterilizáló anyagokra: olajok, porok, éles eszközök és üvegedények. Száraz hőt vagy termikus inaktiválást-sterilizálást alkalmaznak alternatív módszerként a fertőző hulladékok kezelésére.
A pasztőrözés elpusztítja a kórokozó mikroorganizmusokat azáltal, hogy az anyagot 15 másodpercig gyorsan 71,1 °C-ra melegíti, majd gyorsan lehűti. A pasztőrözés nem sterilizálás, mert nem minden mikroorganizmus érzékeny rá. Ezzel a módszerrel megszüntették az olyan betegségek táplálékátvitelét, mint az emésztőrendszeri tuberkulózis és a Q-láz.
Tindalizáció egy szakaszos hősterilizálási módszer, amellyel az oldatokban lévő összes baktérium elpusztítható. Mivel a szaporodó baktériumok könnyen elpusztulnak rövid forralással (5 alkalommal 1 órán át 5 percig), mindössze annyit kell tenni, hogy hagyjuk az oldatot egy bizonyos ideig állni, mielőtt a hő jelentős veszteséggel megzavarná a spórák érését. hőállóságukról.
Szűrés egy olyan eljárás, amelyet a baktériumok és mikroszkopikus részek eltávolítására használnak oldatokból, levegőből és egyéb gázokból. A szűréssel történő sterilizálás legáltalánosabb alkalmazása a laboratóriumban a diagnosztikai készítmények, táptalajok, szövettenyésztő táptalajok, szérumok és szérumkomponenseket tartalmazó oldatok feldolgozása. A szűrés másik gyakori alkalmazása a levegő és a gázok sterilizálása. A pórusátmérő szerint (kb. 12-0,22 μm) megkülönböztetett műanyag vagy papír membránszűrők, amelyeket mechanikai elválasztásra használnak, szintén arra szolgálnak, hogy a mikrobák folyadékokból gyűjtsék össze mikroszkópos vizsgálathoz vagy tenyésztéshez közvetlenül a szűrőn, amikor azt tápanyaggal impregnált felületre helyezik. közepes.
Ultraibolya besugárzás az elektromágneses hullámsugárzás egy fajtája, amely a sejt nukleinsavakra hat. A mikroorganizmusok nagyon érzékenyek a 254 nm hullámhosszú ultraibolya sugarakra. Az ultraibolya fényt a legszélesebb körben használják a levegőben vagy bármilyen felületen lévő mikroorganizmusok elpusztítására. Egyéb alkalmazások közé tartozik bizonyos vegyszerek és műanyagok gyógyszerészeti célú hidegsterilizálása, sejttenyészetek szérumsterilizálása és vízfertőtlenítés. Az ultraibolya besugárzás jelentős hátránya sterilizálóként, hogy nem képes áthatolni az anyagokon.
Ionizáló sugárzás az elektromágneses spektrumban halálos hatással van a mikroorganizmusokra. Ez a spektrum magában foglalja a mikrohullámokat, a gamma-sugarakat, a röntgensugarakat és az elektronáramlást. Az ionizáló sugárzás halálos hatása a célmolekulára gyakorolt közvetlen hatás miatt következik be, ami energiaátvitelt eredményez a molekulába; és közvetett hatás miatt - a gyökök diffúziója.
Ultrahangos energia alacsony gyakorisággal inaktiválja a mikroorganizmusokat vizes oldatokban. A szonikáció fizikai hatása a kavitáció következtében jelentkezik. Ultrahangos tisztítószereket és egyéb eszközöket gyakran használnak műszerek tisztítására, de nem tekintik sterilizálónak. Az ultrahang és a kémiai kezelés kombinálása azonban elpusztítja a mikroorganizmusokat.
Vegyi sterilizátorok
2 % glutáraldehid Folyékony vegyi sterilizálóként korábban széles körben használták olyan orvosi és sebészeti anyagok feldolgozására, amelyek hővel vagy sugárzással nem sterilizálhatók. A glutáraldehidet vakcinák készítéséhez is használják.
Fertőtlenítés.
A fertőtlenítés kémiai módszerekkel vagy forralással végezhető. A forralás hatékony módszer a berendezések, például tűk és fecskendők fertőtlenítésére, ha nem áll rendelkezésre autokláv. Az előre megtisztított orvosi műszereket 20 percig forralni kell. A kémiai fertőtlenítést olyan hőérzékeny berendezéseknél alkalmazzák, amelyek a magas hőmérséklet miatt károsodhatnak. A kémiai fertőtlenítőszereket, például klórkomponenseket, etil- és izopropil-alkoholt, kvaterner ammóniumkomponenseket és glutáraldehidet széles körben alkalmaznak.
Kémiai fertőtlenítőszerek.
Alkohol (etil- és izopropil), vízben 60-85%-os koncentrációban oldva nagyon hatékony a fertőtlenítésben. Az alkoholok baktericid, gombaölő és tuberkulocid hatásúak, de nem befolyásolják a spórákat. Az etil-alkohol vírusölő aktivitása szélesebb, mint az izopropil-alkohol, így hatékonyabb a lipofil és hidrofil vírusok ellen.
Megoldás 37% formaldeid, amit formalinnak hívnak, sterilizálóként, míg 3-8%-os koncentrációja fertőtlenítőszerként használható.
Fenol tiszta formájában nem használják fertőtlenítőszerként toxicitása, daganatok kialakulását és korróziót kiváltó képessége miatt. Fertőtlenítőszerként széles körben alkalmazzák azokat a fenolszármazékokat, amelyekben egy funkciós csoport (klór, bróm, alkil, benzil, fenil, amil) helyettesíti az aromás gyűrű egyik hidrogénatomját. Az ilyen helyettesítés csökkenti a fenol hátrányait. A fenol komponensek elpusztítják a mikrobákat az enzimrendszerek inaktiválásával, a fehérjék kicsapódásával és a sejtfal és a membrán felbomlásával. Jellemzően 2-5%-os koncentrációkat használnak, az alacsonyabb koncentrációk pedig hosszabb expozíciós időt igényelnek.
Halogének. A laboratóriumi gyakorlatban fertőtlenítésre csak klórt és jódot használnak. Mivel a klór erős oxidálószer, úgy gondolják, hogy az oxidáció révén elpusztítja a mikrobákat. Úgy gondolják, hogy a jód az aminosavak N-H és S-H csoportjaival, valamint a tirozin aminosav fenolos csoportjával és a telítetlen zsírsavak szén-szén kettős kötéseivel reagálva elpusztítja a mikroorganizmusokat. A hagyományos kezelés során 2-5%-os formaldehid oldatot permeteznek gőz jelenlétében 60-80°C hőmérsékleten.
Fertőtlenítő.
Antiszeptikumok a mikrobiológiai laboratóriumokban találhatók, elsősorban a kézmosáshoz használt anyagokban. Ha az egészségügyi személyzet sürgősségi ellátást nyújt a betegeknek antibakteriális szereket tartalmazó anyagokkal, ez csökkenti a kórházban szerzett fertőzések számát. A kézmosószerekben előforduló leggyakoribb vegyszerek az alkoholok, a klórhexidin-glükonát, a jodoforok, a klóroxilenol és a triklozán.
A hulladék és a szemét kezelésének hagyományos módszerei az égetés és a gőzsterilizálás.
Égő a választott módszer a hulladékok és törmelékek feldolgozására. Ez a módszer a hulladékot nem fertőzővé teszi, és megváltoztatja alakját és méretét is. A sterilizálás hatékony módszer a hulladék kezelésére, de nem változtatja meg az alakját. Az autoklávban 121°C-on, legalább 15 percig tartó gőzsterilizálás elpusztítja a mikrobiális élet minden formáját, beleértve a nagyszámú baktériumspórát is. Ez a fajta teljes sterilizálás száraz hővel is elvégezhető 160-170°C hőmérsékleten 2-4 órán keresztül. Ügyeljen azonban arra, hogy a száraz hő érintkezzen a sterilizálandó anyaggal. Ezért a folyadékot tartalmazó palackokat lazán le kell zárni dugókkal vagy vattacsomóval, hogy a gőz és a hő kicserélődhessen a palackokban lévő levegővel. A hulladékot tartalmazó biológiailag veszélyes tartályokat szorosan le kell zárni. A sterilizált biológiailag veszélyes anyagokat megfelelő címkével ellátott tartályokba kell zárni.
Gőzsterilizálás (autokláv). A fertőző törmelék akkor tekinthető dekontamináltnak, ha a vegetatív baktériumok, gombák, mikobaktériumok és lipideket tartalmazó vírusok száma 6 lg-szorosára, a bakteriális endospórák száma pedig 4 lg-szorosára csökken.
A baktériumok táplálkozása.
Táplálás. A baktériumsejt táplálkozásán a képlékeny anyag és energia felszívódásának és asszimilációjának folyamatát kell érteni az átalakulási reakciók eredményeként. . A prokarióták táplálkozási típusai összetettek és változatosak. Különböznek a tápanyagoknak a baktériumsejtbe való bejutásának módjától, a szén- és nitrogénforrásoktól, az energiaszerzés módjától és az elektrondonorok természetétől függően.
A tápanyagok szállítása a sejtbe 3 mechanizmussal hajtható végre: passzív diffúzió, könnyített diffúzió és aktív transzport.
Passzív diffúzió egy nem specifikus energiafüggő folyamat, amely az anyagok koncentráció-gradiense mentén megy végbe (egy nagyobb koncentrációjú közegből az ozmózis törvényei szerint passzívan lép be egy kisebb koncentrációjú közegbe). Passzív diffúzióval korlátozott mennyiségű anyag, egyes ionok és monoszacharidok jutnak be a baktériumsejtbe. Az anyagátvitel sebessége a passzív diffúzió során jelentéktelen, és a szállított molekulák lipofilitásától és méretétől függ.
Könnyített diffúzió anyagok energiafüggetlen transzportja koncentrációgradiens mentén permeáz enzimek segítségével. A permeázok specifikus membránfehérjék, amelyek megkönnyítik az anyagok átjutását a citoplazmatikus membránon. A permeáz magára rögzíti a szállított anyag egy molekuláját, amellyel együtt átjut a citoplazma membránon, majd az „anyag-permeáz” komplex disszociál. A felszabaduló permeázt más molekulák hordozására használják. A prokariótákban csak a glicerin jut a sejtbe megkönnyített diffúzióval. Ebben az esetben a glicerin intracelluláris koncentrációja megfelel a sejten kívülinek. A könnyített diffúzió leginkább az eukarióta mikroorganizmusokra jellemző.
Aktiv szállitás- Ez az anyagok energiafüggő átvitele a sejtbe koncentrációgradiens ellenében, specifikus enzimek segítségével. Az anyagok túlnyomó többsége (ionok, szénhidrátok, aminosavak, lipidek stb.) aktív transzport útján jut be a baktériumsejtbe. Aktív szállítás történhet: (1) a szállított anyag kémiai módosítása nélkül; (2) kémiai módosítással.
Milyen baktériumok léteznek: a baktériumok típusai, osztályozásuk
A baktériumok apró mikroorganizmusok, amelyek sok ezer évvel ezelőtt jelentek meg. A mikrobákat nem lehet szabad szemmel látni, de nem szabad megfeledkezni a létezésükről. Hatalmas számú bacilus létezik. A mikrobiológia tudománya ezek osztályozásával, tanulmányozásával, fajtáival, szerkezeti jellemzőivel és élettanával foglalkozik.
A mikroorganizmusokat tevékenységük típusától és funkciójuktól függően eltérően nevezik. Mikroszkóp alatt megfigyelheti, hogyan lépnek kapcsolatba ezek a kis lények egymással. Az első mikroorganizmusok meglehetősen primitív formájúak voltak, de fontosságukat semmi esetre sem szabad alábecsülni. A bacilusok kezdettől fogva fejlődtek, kolóniákat hoztak létre, és megpróbáltak túlélni a változó éghajlati viszonyok között. A különböző vibriók képesek aminosavak cseréjére a normális növekedés és fejlődés érdekében.
Ma nehéz megmondani, hogy ezeknek a mikroorganizmusoknak hány faja van a Földön (ez a szám meghaladja az egymilliót), de a leghíresebbeket és nevüket szinte minden ember ismeri. Nem számít, milyen mikrobák vannak, vagy hogy hívják őket, mindegyiknek van egy előnye - kolóniákban élnek, ami sokkal könnyebbé teszi számukra az alkalmazkodást és a túlélést.
Először is nézzük meg, milyen mikroorganizmusok léteznek. A legegyszerűbb osztályozás jó és rossz. Más szóval, azok, amelyek károsak az emberi szervezetre, számos betegséget okoznak, és azok, amelyek hasznosak. Ezután részletesen beszélünk arról, hogy melyek a fő hasznos baktériumok, és ismertetjük őket.

A mikroorganizmusokat alakjuk és jellemzőik alapján is osztályozhatja. Valószínűleg sokan emlékeznek arra, hogy az iskolai tankönyvekben egy speciális táblázat volt, amely különféle mikroorganizmusokat ábrázolt, és mellettük volt a jelentés és a természetben betöltött szerepük. Többféle baktérium létezik:
- cocci - kis golyók, amelyek láncra hasonlítanak, mivel egymás után helyezkednek el;
- rúd alakú;
- spirilla, spirocheták (tekervény alakúak);
- vibrios.
Különböző formájú baktériumok
Korábban már említettük, hogy az egyik osztályozás a mikrobákat formájuk szerint típusokra osztja.
A Bacillus baktériumoknak is van néhány jellemzője. Léteznek például rúd alakú típusok hegyes rúddal, vastagított, lekerekített vagy egyenes végű. A rúd alakú mikrobák általában nagyon különbözőek, és mindig káoszban vannak, nem sorakoznak láncba (a streptobacillusok kivételével), és nem kötődnek egymáshoz (kivéve a diplobacillusokat).
A mikrobiológusok közé tartoznak a streptococcusok, a staphylococcusok, a diplococcusok és a gonococcusok a gömb alakú mikroorganizmusok közül. Ezek lehetnek golyópárok vagy hosszú láncok.
Az ívelt bacillusok a spirilla, a spirocheták. Mindig aktívak, de nem termelnek spórákat. A Spirilla biztonságos emberek és állatok számára. Megkülönböztetheti a spirillát a spirochetáktól, ha odafigyel a tekercsek számára, amelyek kevésbé csavarodtak, és a végtagjaikon speciális zászlók találhatók.
A patogén baktériumok típusai
Például a coccusnak nevezett mikroorganizmusok egy csoportja, pontosabban a streptococcusok és a staphylococcusok valódi gennyes betegségek (furunculosis, streptococcus mandulagyulladás) okozóivá válnak.
Az anaerobok jól élnek és fejlődnek oxigén nélkül is, bizonyos típusú mikroorganizmusok számára az oxigén végzetes lesz. Az aerob mikrobák fejlődéséhez oxigénre van szükség.
Az archaeák gyakorlatilag színtelen egysejtű szervezetek.
Óvakodni kell a kórokozó baktériumoktól, mert fertőzéseket okoznak, a gram-negatív mikroorganizmusok ellenállnak az antitesteknek. Nagyon sok információ áll rendelkezésre a talajról, a rothadó mikroorganizmusokról, amelyek károsak vagy hasznosak lehetnek.

Általánosságban elmondható, hogy a spirilla nem veszélyes, de egyes fajok sodokut okozhatnak.
A jótékony baktériumok típusai
Még az iskolások is tudják, hogy a bacilusok hasznosak és károsak lehetnek. Az emberek fülről ismernek néhány nevet (staphylococcus, streptococcus, pestisbacilus). Ezek olyan káros lények, amelyek nem csak a külső környezetbe, hanem az emberekbe is beleavatkoznak. Vannak mikroszkopikus bacilusok, amelyek ételmérgezést okoznak.
Mindenképpen hasznos információkat kell tudnia a tejsavról, az élelmiszerekről és a probiotikus mikroorganizmusokról. Például a probiotikumokat, más szóval jó szervezeteket gyakran használják gyógyászati célokra. Kérdezhetitek: minek? Nem engedik, hogy az emberben elszaporodjanak a káros baktériumok, erősítik a belek védő funkcióit, jó hatással vannak az emberi immunrendszerre.
A bifidobaktériumok szintén nagyon jótékony hatással vannak a belekre. A tejsavvibriók körülbelül 25 fajt tartalmaznak. Hatalmas mennyiségben megtalálhatók az emberi szervezetben, de nem veszélyesek. Éppen ellenkezőleg, megvédik a gyomor-bélrendszert a rothadó és más mikrobáktól.
Ha már a jókról beszélünk, nem szabad megemlíteni a streptomycetes hatalmas fajait. Ismerik azokat, akik kloramfenikolt, eritromicint és hasonló gyógyszereket szedtek.
Vannak mikroorganizmusok, mint például az Azotobacter. Hosszú évekig élnek a talajban, jótékony hatással vannak a talajra, serkentik a növények növekedését, megtisztítják a talajt a nehézfémektől. Nélkülözhetetlenek az orvostudományban, a mezőgazdaságban, az orvostudományban és az élelmiszeriparban.
A bakteriális variabilitás típusai
Természetüknél fogva a mikrobák nagyon ingadozóak, gyorsan elpusztulnak, lehetnek spontánok vagy indukáltak. A baktériumok variabilitását nem részletezzük, mivel ez az információ sokkal érdekesebb azoknak, akik érdeklődnek a mikrobiológia és annak minden ága iránt.
Baktériumok típusai szeptikus tartályokhoz
A magánházak lakói megértik, hogy sürgősen meg kell tisztítani a szennyvizet, valamint a szemétmedencéket. Ma már gyorsan és hatékonyan tisztíthatja a lefolyókat a szeptikus tartályok speciális baktériumaival. Ez óriási megkönnyebbülés az ember számára, hiszen a csatornatisztítás nem kellemes feladat.
Már tisztáztuk, hol alkalmazzák a biológiai szennyvízkezelést, most pedig beszéljünk magáról a rendszerről. A szeptikus tartályok baktériumait laboratóriumban termesztik, elpusztítják a szennyvíz kellemetlen szagát, fertőtlenítik a vízelvezető kutakat, a csatornákat, csökkentik a szennyvíz mennyiségét. Háromféle baktériumot használnak szeptikus tartályokhoz:
- aerobic;
- anaerob;
- élő (bioaktivátorok).

Nagyon gyakran az emberek kombinált tisztítási módszereket alkalmaznak. Szigorúan kövesse a terméken található utasításokat, biztosítva, hogy a vízszint elősegítse a baktériumok normális túlélését. Ne felejtse el legalább kéthetente egyszer használni a lefolyót, hogy enni adjon a baktériumoknak, különben elpusztulnak. Ne felejtse el, hogy a tisztítóporokból és folyadékokból származó klór elpusztítja a baktériumokat.
A legnépszerűbb baktériumok a Doctor Robic, a Septifos, a Waste Treat.
A vizeletben lévő baktériumok típusai
Elméletileg a vizeletben nem lehet baktérium, de különféle akciók és helyzetek után az apró mikroorganizmusok ott telepednek meg, ahol akarnak: a hüvelyben, az orrban, a vízben stb. Ha a tesztek során baktériumokat észlelnek, ez azt jelenti, hogy a személy vese-, húgyhólyag- vagy húgycsőbetegségben szenved. A mikroorganizmusok többféle módon is bejuthatnak a vizeletbe. A kezelés előtt nagyon fontos megvizsgálni és pontosan meghatározni a baktériumok típusát és a bejutási utat. Ezt biológiai vizelettenyésztéssel lehet meghatározni, amikor a baktériumok kedvező élőhelyre kerülnek. Ezután ellenőrizzük a baktériumok reakcióját különböző antibiotikumokra.
Kívánjuk, hogy mindig egészséges maradjon. Vigyázz magadra, moss rendszeresen kezet, óvd szervezetedet a káros baktériumoktól!
2.1. A mikrobák rendszertana és nómenklatúrája
A mikrobiális világ sejtes és nem sejtes formákra osztható. A mikrobák sejtes formáit baktériumok, gombák és protozoonok képviselik. Ezeket mikroorganizmusoknak nevezhetjük. A nem sejtes formákat vírusok, viroidok és prionok képviselik.
A sejtes mikrobák új osztályozása a következő taxonómiai egységeket tartalmazza: tartományok, királyságok, típusok, osztályok, rendek, családok, nemzetségek, fajok. A mikroorganizmusok osztályozása genetikai rokonságukon, valamint morfológiai, fiziológiai, antigén- és molekuláris biológiai tulajdonságaikon alapul.
A vírusokat gyakran nem élőlényeknek, hanem autonóm genetikai struktúráknak tekintik, ezért külön-külön vizsgáljuk őket.
A mikrobák sejtes formáit három tartományra osztják. Domains BaktériumokÉs Archaebacteriumok prokarióta típusú sejtszerkezetű mikrobákat tartalmaznak. Domain képviselői Eukarya eukarióták. 4 királyságból áll:
Gomba királyságok (Gomák, Eumycota);
protozoa birodalmai (protozoonok);
királyságok Chromista(króm);
Nem meghatározott taxonómiai helyzetű mikrobák (Microspora, microsporidia).
A prokarióta és eukarióta sejtek szerveződésében mutatkozó különbségeket a táblázat mutatja be. 2.1.
2.1. táblázat. Prokarióta és eukarióta sejt jelei
2.2. A baktériumok osztályozása és morfológiája
A "baktérium" kifejezés a szóból származik baktériumok, mit jelent a bot? A baktériumok prokarióták. Két tartományra oszthatók: BaktériumokÉs Archaebacteriumok. A tartományba tartozó baktériumok archaebaktériumok, az élet egyik legrégebbi formáját képviselik. A sejtfal szerkezeti jellemzői (hiányzik belőlük a peptidoglikán) és a riboszomális RNS. Köztük nincs fertőző betegségek kórokozója.
Egy tartományon belül a baktériumokat a következő taxonómiai kategóriákra osztják: osztály, törzs, rend, család, nemzetség, faj. Az egyik fő taxonómiai kategória az faj. A faj azonos eredetű és genotípusú egyedek gyűjteménye, amelyeket hasonló tulajdonságok egyesítenek, amelyek megkülönböztetik őket a nemzetség többi képviselőjétől. A fajnév bináris nómenklatúrának felel meg, azaz. két szóból áll. Például a diftéria kórokozóját így írják Corynebacterium diphtheriae. Az első szó a nemzetség neve, és nagybetűvel írják, a második szó a fajt jelöli, és kisbetűvel írják.
Egy faj újbóli említésekor a generikus név kezdőbetűjére rövidül, pl. C. diftéria.
Tápközegen izolált homogén mikroorganizmusok halmazát, amelyet hasonló morfológiai, színezési (festékekkel kapcsolatos), kulturális, biokémiai és antigén tulajdonságokkal jellemeznek, ún. tiszta kultúra. Egy meghatározott forrásból izolált és a faj többi tagjától eltérő mikroorganizmusok tiszta kultúráját nevezzük törzs. A „törzs” fogalmához közel áll a „klón” fogalma. A klón egyetlen mikrobiális sejtből származó leszármazottak gyűjteménye.
A mikroorganizmusok bizonyos csoportjainak megjelölésére, amelyek bizonyos tulajdonságokban különböznek egymástól, a „var” (fajta) utótagot használjuk, ezért a mikroorganizmusokat a különbségek természetétől függően morfovaroknak (morfológiai különbség), rezisztens termékeknek (rezisztenciakülönbség) jelöljük. például antibiotikumokra), szerovarokra (az antigének különbsége), fagovarokra (a bakteriofágokkal szembeni érzékenység különbsége), biovarokra (a biológiai tulajdonságok különbsége), a kemovarokra (a biokémiai tulajdonságok különbsége) stb.
Korábban a baktériumok osztályozásának alapja a sejtfal szerkezeti jellemzője volt. A baktériumok sejtfal szerkezeti sajátosságai szerinti osztódása a Gram-módszerrel színezésük lehetséges változatosságával függ össze. E módszer szerint, amelyet H. Gram dán tudós 1884-ben javasolt, a festési eredményektől függően a baktériumokat Gram-pozitív, kék-lilára festett és Gram-negatív, vörösre festett baktériumokra osztják.
Jelenleg a besorolás a genetikai rokonság fokán alapul, a riboszomális RNS (rRNS) genom szerkezetének vizsgálatán (lásd 5. fejezet), a genomban található guanin citozin párok (GC párok) százalékos meghatározásán, egy A genom restrikciós térképe, valamint a hibridizáció mértékének tanulmányozása. Figyelembe veszik a fenotípusos mutatókat is: a Gram-festéshez való hozzáállást, a morfológiai, kulturális és biokémiai tulajdonságokat, az antigén szerkezetet.
Tartomány Baktériumok 23 típust foglal magában, amelyek közül az alábbiak orvosi jelentőségűek.
A legtöbb gram-negatív baktérium a törzsbe csoportosul Proteobaktériumok(a görög istenről kapta a nevét Proteus, különböző formákat ölteni képes). típus Proteobaktériumok 5 osztályra osztva:
Osztály Alfaproteobaktériumok(születés Rickettsia, Orientia, Erlichia, Bartonella, Brucella);
Osztály Betaproteobaktériumok(születés Bordetella, Burholderia, Neisseria, Spirillum);
Osztály Gammaproteobaktériumok(a család képviselői Enterobacteriaceae szülés Francisella, Legionella, Coxiella, Pseudomonas, Vibrio);
Osztály Deltaproteobaktériumok(nemzetség bilophila);
Osztály Epszilonproteobaktériumok(születés Campilobacter, Helicobacter). A Gram-negatív baktériumok közé tartoznak a következő típusok is:
típus Chlamydiae(születés Chlamydia, Chlamydophila), típus Spirocheták(születés Spirocheta, Borrelia, Treponema, Leptospira); típus Bacteroides(születés Bacteroides, Prevotella, Porphyromonas).
A Gram-pozitív baktériumok a következő típusúak:
típus Cégek osztályt tartalmazza Clostridium(születés Clostridium, Peptococcus), Osztály Bacillusok (Listeria, Staphylococcus, Lactobacillus, Streptococcus)és osztály Mollicutes(születés Mycoplasma, Ureaplasma), amelyek olyan baktériumok, amelyeknek nincs sejtfaluk;
típus Actinobacteriumok(születés Actinomyces, Micrococcus, Corynebacterium, Mycobacterium, Gardnerella, Bifidobacterium, Propionibacterium, Mobiluncus).
2.2.1. A baktériumok morfológiai formái
A baktériumoknak több fő formája létezik: kokcoid, rúd alakú, tekercses és elágazó (2.1. ábra).
Gömb alakú formák, vagy coccusok- 0,5-1 mikron méretű gömbbaktériumok, amelyek relatív helyzetük szerint mikrococcusokra, diplococcusokra, streptococcusokra, tetracoccusokra, sarcinákra és staphylococcusokra oszthatók.
Micrococcusok (görögből. mikros- kicsi) - külön elhelyezkedő sejtek.
Diplococci (görögből. diploos- kettős), vagy páros coccusok, párban helyezkednek el (pneumococcus, gonococcus, meningococcus), mivel a sejtek osztódás után nem válnak el. A Pneumococcus (a tüdőgyulladás kórokozója) lándzsa alakú az ellenkező oldalon, a gonococcus (a gonorrhoea kórokozója) és a meningococcus (a tüdőgyulladás kórokozója)
 Rizs. 2.1. A baktériumok alakjai
Rizs. 2.1. A baktériumok alakjai
járványos agyhártyagyulladás okozója) kávébab alakúak, homorú felületük egymással szemben.
Streptococcusok (görögből. streptos- lánc) - kerek vagy hosszúkás alakú sejtek, amelyek láncot alkotnak a sejtosztódás miatt ugyanabban a síkban, és fenntartják közöttük a kapcsolatot az osztódás helyén.
Sarcins (a lat. sarcina- csomó, bála) 8 vagy több kókuszból álló csomagok formájában vannak elrendezve, mivel a sejtosztódás során három egymásra merőleges síkban jönnek létre.
Staphylococcusok (görögből. stafil- szőlőfürt) - szőlőfürt formájában lévő coccusok a különböző síkok felosztása következtében.
Rúd alakú baktériumok különböznek méretükben, a sejtvégek alakjában és a sejtek egymáshoz viszonyított helyzetében. A sejt hossza 1-10 µm, vastagsága 0,5-2 µm. A botoknak igazuk lehet
(Escherichia coli stb.) és szabálytalan bot alakú (Corynebacteria stb.) alakú. A legkisebb rúd alakú baktériumok közé tartozik a rickettsia.
A rudak végei levághatók (anthrax bacillus), lekerekítettek (Escherichia coli), hegyesek (fuzobaktériumok) vagy megvastagodás formájában. Ez utóbbi esetben a rúd úgy néz ki, mint egy ütő (Corynebacterium diphtheria).
Az enyhén ívelt rudakat vibrióknak (Vibrio cholerae) nevezik. A legtöbb pálcika alakú baktérium véletlenszerűen helyezkedik el, mivel a sejtek osztódás után eltávolodnak egymástól. Ha az osztódás után a sejtek a sejtfal közös töredékeivel összekapcsolva maradnak, és nem térnek el egymástól, akkor egymással szöget zárva helyezkednek el (Corynebacterium diphtheria), vagy láncot alkotnak (antrax bacillus).
Csavart formák- spirál alakú baktériumok, amelyeknek két típusa van: spirilla és spirocheta. A Spirilla dugóhúzó alakú, csavart sejteknek tűnik, nagy fürtökkel. A kórokozó spirillák közé tartozik a sodoku (patkánycsípés betegség), valamint a campylobacter és a helicobacter kórokozója, amelyek görbéi egy repülő sirály szárnyaira emlékeztetnek. A spirocheták vékony, hosszú, tekert baktériumok, amelyek kisebb fürtökben és mozgásmintájukban különböznek a spirillától. Felépítésük sajátosságait az alábbiakban ismertetjük.
Elágazás - A bifidobaktériumokban található Y-alakú ágakkal rendelkező pálcika alakú baktériumok fonalas elágazó sejtek formájában is megjelenhetnek, amelyek összefonódva micéliumot képezhetnek, amint azt az aktinomicétákban megfigyelték.
2.2.2. A bakteriális sejt szerkezete
A baktériumok szerkezetét jól tanulmányozták teljes sejtek és ultravékony metszeteik elektronmikroszkópos vizsgálatával, valamint egyéb módszerekkel. A baktériumsejtet sejtfalból és citoplazmatikus membránból álló membrán veszi körül. A héj alatt protoplazma található, amely zárványokkal ellátott citoplazmából és egy örökletes készülékből áll - a mag analógja, az úgynevezett nukleoid (2.2. ábra). Vannak további struktúrák: kapszula, mikrokapszula, nyálka, flagella, pili. Egyes baktériumok kedvezőtlen körülmények között képesek spórákat képezni.
 Rizs. 2.2. A baktériumsejt felépítése: 1 - kapszula; 2 - sejtfal; 3 - citoplazmatikus membrán; 4 - mezoszómák; 5 - nukleoid; 6 - plazmid; 7 - riboszómák; 8 - zárványok; 9 - flagellum; 10 - pili (villi)
Rizs. 2.2. A baktériumsejt felépítése: 1 - kapszula; 2 - sejtfal; 3 - citoplazmatikus membrán; 4 - mezoszómák; 5 - nukleoid; 6 - plazmid; 7 - riboszómák; 8 - zárványok; 9 - flagellum; 10 - pili (villi)
Sejtfal- erős, rugalmas szerkezet, amely bizonyos formát ad a baktériumnak, és az alatta lévő citoplazmatikus membránnal együtt visszafogja a magas ozmotikus nyomást a baktériumsejtben. Részt vesz a sejtosztódás folyamatában és a metabolitok szállításában, receptorai vannak bakteriofágokhoz, bakteriocinokhoz és különféle anyagokhoz. A legvastagabb sejtfal a gram-pozitív baktériumokban található (2.3. ábra). Tehát, ha a Gram-negatív baktériumok sejtfalának vastagsága körülbelül 15-20 nm, akkor a Gram-pozitív baktériumokban elérheti az 50 nm-t vagy még többet is.
A bakteriális sejtfal alapja az peptidoglikán. A peptidoglikán egy polimer. Párhuzamos poliszacharid-glikánláncok képviselik, amelyek ismétlődő N-acetil-glükózamin és N-acetil-muraminsav-maradékokból állnak, amelyeket glikozidos kötéssel kapcsolnak össze. Ezt a kötést megszakítja a lizozim, amely egy acetil-muramidáz.
Egy tetrapeptid kovalens kötéssel kapcsolódik az N-acetil-muraminsavhoz. A tetrapeptid L-alaninból áll, amely N-acetil-muraminsavhoz kapcsolódik; D-glutamin, amely a gram-pozitív baktériumokban L-lizinnel, a gram-tri-
 Rizs. 2.3. A bakteriális sejtfal felépítésének vázlata
Rizs. 2.3. A bakteriális sejtfal felépítésének vázlata
hasznos baktériumok - diaminopimelinsavval (DAP), amely a lizin prekurzora az aminosavak bakteriális bioszintézisének folyamatában, és egyedülálló vegyület, amely csak baktériumokban van jelen; A 4. aminosav a D-alanin (2.4. ábra).
A gram-pozitív baktériumok sejtfala kis mennyiségben poliszacharidokat, lipideket és fehérjéket tartalmaz. E baktériumok sejtfalának fő összetevője a többrétegű peptidoglikán (murein, mukopeptid), amely a sejtfal tömegének 40-90%-át teszi ki. A Gram-pozitív baktériumok peptidoglikán különböző rétegeinek tetrapeptidjei 5 glicin-maradékból álló polipeptidláncokkal (pentaglicin) kapcsolódnak egymáshoz, ami merev geometriai szerkezetet ad a peptidoglikánnak (2.4. ábra, b). Kovalensen kapcsolódik a gram-pozitív baktériumok sejtfalának peptidoglikánjához teichoinsavak(görögből tekhos- fal), melynek molekulái 8-50 glicerin- és ribitolmaradékból álló láncok, amelyeket foszfáthidak kötnek össze. A baktériumok alakját és erejét a többrétegű peptidoglikán merev rostos szerkezete adja, peptidek keresztkötéseivel.
 Rizs. 2.4. A peptidoglikán szerkezete: a - Gram-negatív baktériumok; b - Gram-pozitív baktériumok
Rizs. 2.4. A peptidoglikán szerkezete: a - Gram-negatív baktériumok; b - Gram-pozitív baktériumok
A Gram-pozitív baktériumok azon képessége, hogy megőrizzék az enciánibolyát jóddal kombinálva, ha Gram-festéssel (a baktériumok kék-lila színe) festenek, a többrétegű peptidoglikánnak a festékkel való kölcsönhatási tulajdonságával függ össze. Ezenkívül a bakteriális kenet utólagos alkoholos kezelése a peptidoglikán pórusainak szűkülését okozza, és ezáltal megtartja a festéket a sejtfalban.
A Gram-negatív baktériumok alkohollal való érintkezés után elveszítik a festéket, ami a kisebb mennyiségű peptidoglikánnak köszönhető (a sejtfal tömegének 5-10%-a); alkohollal elszíneződnek, fukszinnal vagy szafraninnal kezelve kipirosodnak. Ez a sejtfal szerkezeti sajátosságainak köszönhető. A Gram-negatív baktériumok sejtfalában a peptidoglikánt 1-2 réteg képviseli. A rétegek tetrapeptidjei közvetlen peptidkötéssel kapcsolódnak egymáshoz az egyik tetrapeptid DAP aminocsoportja és egy másik réteg tetrapeptidjének D-alanin karboxilcsoportja között (2.4. ábra, a). A peptidoglikánon kívül van egy réteg lipoprotein, a DAP-n keresztül kapcsolódik a peptidoglikánhoz. Következett külső membrán sejtfal.
Külső membrán egy mozaik szerkezet, amely lipopoliszacharidokból (LPS), foszfolipidekből és fehérjékből áll. Belső rétegét foszfolipidek képviselik, a külső rétegben LPS található (2.5. ábra). Így a külső mem-
 Rizs. 2.5. Lipopoliszacharid szerkezet
Rizs. 2.5. Lipopoliszacharid szerkezet
a brán aszimmetrikus. Az LPS külső membrán három részből áll:
A lipid A konzervatív szerkezetű, majdnem ugyanaz a Gram-negatív baktériumokban. A lipid A foszforilált glükózamin diszacharid egységekből áll, amelyekhez hosszú zsírsavláncok kapcsolódnak (lásd a 2.5. ábrát);
Mag, vagy mag, kéregrész (a lat. mag- mag), viszonylag konzervatív oligoszacharid szerkezet;
Nagyon variábilis O-specifikus poliszacharid lánc, amely azonos oligoszacharid szekvenciák ismétlődéséből jön létre.
Az LPS-t a lipid A rögzíti a külső membránban, ami LPS toxicitást okoz, ezért endotoxinnal azonosítják. A baktériumok antibiotikumok általi elpusztítása nagy mennyiségű endotoxin felszabadulásához vezet, ami endotoxikus sokkot okozhat a betegben. Az LPS magja vagy magrésze a lipid A-ból nyúlik ki. Az LPS mag legállandóbb része a ketodezoxioktonsav. O-specifikus poliszacharid lánc, amely az LPS molekula magjából nyúlik ki,
ismétlődő oligoszacharid egységekből áll, meghatározza egy adott baktériumtörzs szerocsoportját, szerovariációját (az immunszérum segítségével kimutatható baktériumtípus). Így az LPS fogalma összefügg az O-antigén fogalmával, amellyel a baktériumok megkülönböztethetők. A genetikai változások defektusokhoz, a bakteriális LPS lerövidüléséhez vezethetnek, és ennek eredményeként az R-formák durva kolóniáinak megjelenéséhez, amelyek elveszítik O-antigén-specifitását.
Nem minden gram-negatív baktérium rendelkezik teljes O-specifikus poliszacharid lánccal, amely ismétlődő oligoszacharid egységekből áll. Különösen a nemzetséghez tartozó baktériumok Neisseria rövid glikolipidje van, amelyet lipooligoszacharidnak (LOS) neveznek. Összehasonlítható az R formával, amely elvesztette O-antigén specifitását, amit a durva mutáns törzsekben figyeltek meg E. coli. A VOC szerkezete hasonlít a humán citoplazma membrán glikoszfingolipidjének szerkezetére, így a VOC utánozza a mikrobát, lehetővé téve az számára, hogy elkerülje a gazdaszervezet immunválaszát.
A külső membrán mátrixfehérjéi úgy hatolnak át rajta, hogy a fehérjemolekulák ún porinami, határos hidrofil pórusok, amelyeken víz és kis hidrofil molekulák haladnak át, amelyek relatív tömege legfeljebb 700 D.
A külső és a citoplazmatikus membrán között van periplazmatikus tér, vagy enzimeket (proteázokat, lipázokat, foszfatázokat, nukleázokat, β-laktamázokat) tartalmazó periplazmák, valamint a transzportrendszerek komponensei.
Ha a bakteriális sejtfal szintézise a lizozim, a penicillin, a test védőfaktorai és más vegyületek hatására megszakad, megváltozott (gyakran gömb alakú) sejtek képződnek: protoplasztok- sejtfalat teljesen hiányzó baktériumok; szferoplasztok- részben megőrzött sejtfalú baktériumok. A sejtfal-inhibitor eltávolítása után az ilyen módosult baktériumok visszafordíthatók, pl. teljes értékű sejtfalat szerezzen és visszaállítsa eredeti formáját.
Azokat a szferoid vagy protoplaszt típusú baktériumokat, amelyek antibiotikumok vagy más tényezők hatására elvesztették a peptidoglikán szintetizálásának képességét, és képesek szaporodni, ún. L-alakúak(a D. Lister Intézet nevéből, ahol először
tanulmányozták). Az L-formák mutációk következtében is kialakulhatnak. Ozmózisérzékeny, gömb alakú, különböző méretű lombik alakú sejtek, beleértve a baktériumszűrőkön áthaladókat is. Egyes L-formák (instabilok), ha a baktériumok változását okozó tényezőt eltávolítják, megfordulhatnak, és visszatérnek az eredeti baktériumsejtbe. L-formákat számos fertőző betegség kórokozója képes előállítani.
Citoplazma membrán ultravékony metszetek elektronmikroszkópos vizsgálatánál háromrétegű membrán (2 sötét réteg, egyenként 2,5 nm vastag, egy világos köztes réteg választja el). Szerkezetében hasonló az állati sejtek plazmalemmájához, és kettős lipidrétegből, főleg foszfolipidekből áll, beágyazott felülettel és integrált fehérjékkel, amelyek úgy tűnik, hogy áthatolnak a membrán szerkezetén. Ezek egy része az anyagok szállításában részt vevő permeáz. Az eukarióta sejtekkel ellentétben a baktériumsejt citoplazmatikus membránjában hiányoznak a szterolok (a mikoplazmák kivételével).
A citoplazmatikus membrán egy dinamikus szerkezet mobil komponensekkel, ezért úgy gondolják, mint egy mozgó folyadékszerkezetet. Körülveszi a baktériumok citoplazmájának külső részét, és részt vesz az ozmotikus nyomás szabályozásában, az anyagok szállításában és a sejt energia-anyagcseréjében (az elektrontranszport lánc enzimei, az adenozin-trifoszfatáz - ATPáz stb. miatt). A túlzott növekedéssel (a sejtfal növekedéséhez képest) a citoplazma membrán invaginátumok - invaginációk bonyolultan csavarodott membránszerkezetek formájában, ún. mezoszómák. A kevésbé bonyolultan csavart struktúrákat intracitoplazmatikus membránoknak nevezzük. A mezoszómák és az intracitoplazmatikus membránok szerepe nem teljesen ismert. Még azt is feltételezik, hogy olyan műtermékről van szó, amely az elektronmikroszkópos minta előkészítése (rögzítése) után keletkezik. Ennek ellenére úgy gondolják, hogy a citoplazma membrán származékai részt vesznek a sejtosztódásban, energiát adva a sejtfal szintéziséhez, és részt vesznek az anyagok kiválasztásában, a sporulációban, pl. nagy energiafogyasztású folyamatokban. A citoplazma a baktériumok fő térfogatát foglalja el
sejt, és oldható fehérjékből, ribonukleinsavakból, zárványokból és számos kis szemcsékből - riboszómákból áll, amelyek a fehérjék szintéziséért (transzlációjáért) felelősek.
Riboszómák A baktériumok mérete körülbelül 20 nm, ülepedési együtthatója pedig 70S, ellentétben az eukarióta sejtekre jellemző 80S riboszómákkal. Ezért egyes antibiotikumok a bakteriális riboszómákhoz való kötődés révén gátolják a bakteriális fehérjeszintézist anélkül, hogy befolyásolnák az eukarióta sejtekben a fehérjeszintézist. A bakteriális riboszómák két alegységre disszociálhatnak: 50S és 30S. Az rRNS a baktériumok konzervált eleme (az evolúció „molekuláris órája”). A 16S rRNS a kis riboszomális alegység, a 23S rRNS pedig a nagy riboszomális alegység része. A 16S rRNS vizsgálata a génszisztematika alapja, lehetővé téve az élőlények rokonsági fokának felmérését.
A citoplazma különféle zárványokat tartalmaz glikogén granulátum, poliszacharidok, β-hidroxi-vajsav és polifoszfátok (volutin) formájában. Felhalmozódnak, ha a környezetben túl sok tápanyag van, és tartalék anyagként szolgálnak a táplálkozáshoz és az energiaszükséglethez.
Volyutin affinitása van az alapfestékekhez, és speciális festési módszerekkel (például Neisser szerint) könnyen kimutatható metakromatikus granulátum formájában. A toluidin kékkel vagy metilénkékkel a volutin vörös-ibolyára festődik, a baktérium citoplazmája pedig kékre festődik. A diftéria bacillusban a volutin szemcsék jellegzetes elrendeződése intenzíven festődő sejtpólusok formájában mutatkozik meg. A volutin metakromatikus elszíneződése a polimerizált szervetlen polifoszfát magas tartalmához kapcsolódik. Elektronmikroszkóp alatt 0,1-1 mikron méretű elektronsűrű szemcséknek tűnnek.
Nukleoid- egyenértékű a baktériumok sejtmagjával. A baktériumok központi zónájában található, kétszálú DNS formájában, szorosan összetömörítve, mint egy labda. A baktériumok nukleoidja az eukariótáktól eltérően nem rendelkezik magburokkal, nukleolusszal és bázikus fehérjékkel (hisztonokkal). A legtöbb baktérium egy kromoszómát tartalmaz, amelyet egy gyűrűbe zárt DNS-molekula képvisel. De néhány baktériumnak két gyűrű alakú kromoszómája van (V. cholerae)és lineáris kromoszómák (lásd 5.1.1. szakasz). A nukleoid fénymikroszkóppal derül ki DNS-specifikus festéssel történő festés után
módszerek: Feulgen vagy Romanovsky-Giemsa szerint. A baktériumok ultravékony metszeteinek elektrondiffrakciós mintázataiban a nukleoid világos zónákként jelenik meg, a DNS fibrilláris, fonalszerű struktúráival, amelyek bizonyos területeken a citoplazmatikus membránhoz vagy a kromoszóma replikációjában részt vevő mezoszómához kötődnek.
A nukleoidon kívül a baktériumsejt extrakromoszómális öröklődési faktorokat – plazmidokat – tartalmaz (lásd 5.1.2. fejezet), amelyek a DNS kovalensen zárt gyűrűi.
Kapszula, mikrokapszula, nyálka.kapszula - 0,2 mikronnál vastagabb nyálkahártya szerkezet, amely szilárdan kapcsolódik a bakteriális sejtfalhoz, és világosan meghatározott külső határokkal rendelkezik. A kapszula patológiás anyagból származó lenyomatkenetekben látható. A tiszta baktériumtenyészetekben a kapszula ritkábban képződik. Speciális Burri-Gins szerinti kenetfestési módszerekkel mutatják ki, ami negatív kontrasztot hoz létre a kapszula anyagai között: a tinta sötét hátteret hoz létre a kapszula körül. A kapszula poliszacharidokból (exopoliszacharidokból), néha polipeptidekből áll, például az anthrax bacillusban D-glutaminsav polimerjeiből áll. A kapszula hidrofil és nagy mennyiségű vizet tartalmaz. Megakadályozza a baktériumok fagocitózisát. A kapszula antigén: a kapszula elleni antitestek a megnagyobbodást okozzák (kapszuladuzzadási reakció).
Sok baktérium képződik mikrokapszula- 0,2 mikronnál kisebb vastagságú, csak elektronmikroszkóppal kimutatható nyálkahártyaképződés.
Meg kell különböztetni a kapszulától nyálka - nyálkahártya exopoliszacharidok, amelyek nem rendelkeznek világos külső határokkal. A nyálka vízben oldódik.
A mukoid exopoliszacharidok a Pseudomonas aeruginosa nyálkahártya törzseire jellemzőek, gyakran megtalálhatók a cisztás fibrózisban szenvedő betegek köpetében. A bakteriális exopoliszacharidok részt vesznek az adhézióban (az aljzathoz tapadnak); glikokalixnak is nevezik.
A kapszula és a nyálka megvédi a baktériumokat a károsodástól és a kiszáradástól, mivel hidrofil lévén jól megköti a vizet és megakadályozza a makroorganizmus és a bakteriofágok védőfaktorainak hatását.
Flagella baktériumok határozzák meg a baktériumsejt mobilitását. A zászlók vékony szálak, amelyek felvesznek
A citoplazmatikus membránból származnak, és hosszabbak, mint maga a sejt. A flagella vastagsága 12-20 nm, hossza 3-15 µm. Három részből állnak: egy spirálszálból, egy kampóból és egy alaptestből, amely speciális korongokkal ellátott rudat tartalmaz (egy pár korong a gram-pozitív baktériumokban és két pár a gram-negatív baktériumokban). A zászlók a citoplazma membránjához és a sejtfalhoz korongokkal kapcsolódnak. Ez azt a hatást hozza létre, hogy egy villanymotor egy rúddal - egy rotorral - forgatja a flagellumot. A citoplazma membránján lévő protonpotenciál különbséget energiaforrásként használják. A rotációs mechanizmust a proton ATP szintetáz biztosítja. A flagellum forgási sebessége elérheti a 100 rps-t. Ha egy baktériumnak több flagellája van, azok szinkronban forogni kezdenek, egyetlen köteggé fonódnak össze, egyfajta légcsavart alkotva.
A flagellák egy flagellin nevű fehérjéből állnak. (flagellum- flagellum), amely egy antigén - az úgynevezett H-antigén. A flagellin alegységek spirál alakúak.
A különböző baktériumfajokban a flagellák száma egytől (monotrichus) a Vibrio cholerae-ben a baktérium kerülete mentén több tízig és százig terjed (peritrichus), Escherichia coliban, Proteusban stb. A Lophotrich-félék egyik végén egy köteg flagella található a sejt. Az Amphitriciának egy flagellum vagy egy köteg flagella van a sejt ellentétes végein.
A flagellákat nehézfémekkel bevont készítmények elektronmikroszkópos vizsgálatával vagy fénymikroszkóppal mutatják ki, speciális módszerekkel végzett kezelés után, amelyek különböző anyagok maratásán és adszorpcióján alapulnak, ami a flagellák vastagságának növekedéséhez vezet (például ezüstözés után).
Villi vagy pili (fimbriae)- fonalszerű képződmények, vékonyabbak és rövidebbek (3-10 nm * 0,3-10 µm), mint a flagellák. A pilusok a sejtfelszínről nyúlnak ki, és a fehérje pilinből állnak. A pili több fajtája ismert. Az általános típusú pilik felelősek a szubsztrátumhoz való kötődésért, a táplálkozásért és a víz-só anyagcseréért. Számos – sejtenként több száz. A nemi pilusok (sejtenként 1-3) kapcsolatot teremtenek a sejtek között, és a genetikai információkat konjugáció útján továbbítják közöttük (lásd 5. fejezet). Különös érdeklődésre tartanak számot a IV-es típusú pilók, amelyekben a végek hidrofóbok, aminek következtében ezeket a piliket göndörnek is nevezik. Elhelyezkedés
A sejt pólusainál helyezkednek el. Ezek a pilusok patogén baktériumokban találhatók. Antigén tulajdonságokkal rendelkeznek, érintkezésbe hozzák a baktériumokat a gazdasejttel, és részt vesznek a biofilm kialakításában (lásd 3. fejezet). Sok pili a bakteriofágok receptora.
Viták - a nyugvó baktériumok sajátos formája, gram-pozitív típusú sejtfalszerkezettel. A nemzetség spóraképző baktériumai Bacilus, amelyekben a spóra mérete nem haladja meg a sejt átmérőjét, bacilusoknak nevezzük. Azokat a spóraképző baktériumokat, amelyeknél a spóra mérete meghaladja a sejt átmérőjét, így azok orsó alakot vesznek fel, ún. klostridiumok, például a nemzetséghez tartozó baktériumok Clostridium(a lat. Clostridium- orsó). A spórák savállóak, ezért Aujeszky-módszerrel vagy Ziehl-Neelsen-módszerrel vörösre festik, a vegetatív sejtet pedig kékre festik.
A spórák kialakulása, a spórák alakja és elhelyezkedése a sejtben (vegetatív) a baktériumok faji sajátossága, amely lehetővé teszi a baktériumok megkülönböztetését. A spórák alakja lehet ovális vagy gömb alakú, a sejten belüli elhelyezkedésük terminális, azaz. a pálcika végén (a tetanusz kórokozójában), subterminálisan - közelebb a pálcika végéhez (a botulizmus, gáz gangréna kórokozóiban) és központi (az anthrax bacillusban).
A sporuláció (sporuláció) folyamata számos szakaszon megy keresztül, amelyek során a baktérium vegetatív sejt citoplazmájának és kromoszómájának egy része elválik, körülvéve egy benőtt citoplazmatikus membránnal - prospóra képződik.
A prospóra protoplaszt egy nukleoidot, egy fehérjeszintetizáló rendszert és egy glikolízisen alapuló energiatermelő rendszert tartalmaz. A citokrómok még az aerobokban is hiányoznak. Nem tartalmaz ATP-t, a csíráztatáshoz szükséges energiát 3-glicerin-foszfát formájában tárolják.
A prospórát két citoplazmatikus membrán veszi körül. A spóra belső membránját körülvevő réteget ún spórafal, peptidoglikánból áll, és a sejtfal fő forrása a spórák csírázása során.
A külső membrán és a spórafal között vastag réteg képződik, amely peptidoglikánból áll, amely számos keresztkötést tartalmaz - kéreg.
A külső citoplazmatikus membránon kívül található spórahéj, keratinszerű fehérjékből áll,
több intramolekuláris diszulfid kötést tartalmaz. Ez a héj ellenáll a vegyi anyagoknak. Egyes baktériumok spóráinak további burkolata van - exosporium lipoprotein természet. Ily módon többrétegű, rosszul áteresztő héj jön létre.
A sporuláció a prospóra intenzív fogyasztásával, majd a fejlődő dipikolinsav és kalciumionok spórahéjával jár együtt. A spóra hőállóságot szerez, ami a kalcium-dipikolinát jelenlétével függ össze.
A spóra hosszú ideig fennmaradhat a többrétegű héj, a kalcium-dipikolinát, az alacsony víztartalom és a lassú anyagcsere-folyamatok miatt. A talajban például a lépfene és a tetanusz kórokozói évtizedekig fennmaradhatnak.
Kedvező körülmények között a spórák kicsíráznak, és három egymást követő szakaszon mennek keresztül: aktiválás, beindulás, növekedés. Ebben az esetben egy spórából egy baktérium képződik. Az aktiválás készen áll a csírázásra. 60-80 °C hőmérsékleten a spóra aktiválódik a csírázáshoz. A csírázás beindítása néhány percig tart. A kinövési szakaszt gyors növekedés jellemzi, amelyet a héj pusztulása és a palánta kikelése kísér.
2.2.3. A spirocheták, a rickettsia, a chlamydia, az aktinomyceták és a mikoplazmák szerkezeti jellemzői
Spirochetes- vékony, hosszú tekercs baktériumok. Egy külső membrán sejtfalból állnak, amely egy citoplazmatikus hengert vesz körül. A külső membrán tetején glikozaminoglikán jellegű átlátszó borítás található. A sejtfal külső membránja alatt rostok találhatók, amelyek a citoplazmatikus henger körül csavarodnak, így a baktériumok spirális formát kölcsönöznek. A fibrillumok a sejt végeihez kapcsolódnak és egymás felé irányulnak. A fibrillumok száma és elrendezése fajonként változó. A rostok részt vesznek a spirocheták mozgásában, így a sejteknek forgó, hajlító és transzlációs mozgást adnak. Ebben az esetben a spirocheták hurkokat, fürtöket és hajlatokat képeznek, amelyeket másodlagos fürtöknek neveznek. A spirocheták nem érzékelik jól a festékeket. Általában Romanovsky-Giemsa szerint festettek vagy ezüstözöttek. Élő
A spirocheta formáját fáziskontrasztos vagy sötétmezős mikroszkóppal vizsgálják.
A spirochetákat három nemzetség képviseli, amelyek patogén az emberek számára: Treponema, Borrelia, Leptospira.
Treponema(nemzetség Treponema) vékony, dugóhúzóval csavart szálak megjelenése 8-12 egységes kis fürtökkel. A treponema protoplasztja körül 3-4 rost (flagella) található. A citoplazma citoplazmatikus filamentumokat tartalmaz. A kórokozók képviselői T. pallidum- a szifilisz kórokozója, T. pertenue- a trópusi yaws kórokozója. Vannak szaprofiták is - az emberi szájüreg lakói és a tározók iszapja.
Borrelia(nemzetség Borrelia), a treponemákkal ellentétben hosszabbak, 3-8 nagy fürtjük és 7-20 rostjuk van. Ezek közé tartozik a visszaeső láz kórokozója (B. recurrentis)és a Lyme-kór kórokozói (V. burgdorferi)és egyéb betegségek.
Leptospira(nemzetség Leptospira) sekély és gyakori fürtök csavart kötél formájában. Ezeknek a spirochetáknak a végei ívesek, mint a horgok, a végén vastagodással. Másodlagos fürtöket képezve az S vagy C betű alakját veszik fel; két axiális fibrillája van. A kórokozó képviselője L. vallatja vízzel vagy étellel lenyelve leptospirózist okoz, ami vérzést és sárgaságot okoz.
A riketsiák anyagcseréje független a gazdasejttől, azonban lehetséges, hogy a szaporodáshoz nagy energiájú vegyületeket kapnak a gazdasejttől. A kenetekben és a szövetekben Romanovsky-Giemsa, Macchiavello-Zdrodovsky szerint festettek (a rickettsiák vörösek, a fertőzött sejtek kékek).
Emberben a rickettsiák járványos tífuszt okoznak. (R. prowazekii), kullancs által terjesztett rickettsiosis (R. sibirica), Rocky Mountain foltos láz (R. rickettsii)és egyéb rickettsiosisok.
Sejtfaluk szerkezete a gram-negatív baktériumokéhoz hasonlít, bár vannak eltérések. Nem tartalmaz tipikus peptidoglikánt: teljesen hiányzik belőle az N-acetilmuraminsav. A sejtfal kettős külső membránból áll, amely lipopoliszacharidot és fehérjéket tartalmaz. A peptidoglikán hiánya ellenére a chlamydia sejtfala merev. A sejt citoplazmáját a belső citoplazmatikus membrán korlátozza.
A chlamydia kimutatásának fő módszere a Romanovsky-Giemsa festés. A szín az életciklus szakaszától függ: az elemi testek lilának tűnnek a sejt kék citoplazmájának hátterében, a retikuláris testek kéknek.
Emberben a chlamydia károsítja a szemet (trachoma, kötőhártya-gyulladás), az urogenitális traktusban, a tüdőben stb.
Actinomycetes- elágazó, fonalas vagy rúd alakú Gram-pozitív baktériumok. A neve (görögből. actis- Ray, mykes- gomba) kaptak az érintett szövetekben a drusen képződése miatt - szorosan összefonódó szálak szemcséi formájában
a középpontból kinyúló és lombik alakú megvastagodásokban végződő sugarak. Az aktinomyceták a gombákhoz hasonlóan micéliumot - fonalszerűen összefonódó sejteket (hifákat) alkotnak. Szubsztrát micéliumot képeznek, amely a sejt tápközegbe való benövése eredményeként jön létre, és légmicéliumot, amely a táptalaj felületén nő. Az aktinomyceták a micélium feldarabolásával a rúd alakú és coccoid baktériumokhoz hasonló sejtekre osztódhatnak. Az aktinomyceták légi hifáin spórák képződnek, amelyek a szaporodást szolgálják. Az Actinomycete spórái általában nem hőállóak.
Az aktinomicétákkal közös filogenetikai ágat az úgynevezett nocardiform (nocardioform) aktinomicéták alkotják - a szabálytalan alakú, rúd alakú baktériumok kollektív csoportja. Egyéni képviselőik elágazó formákat alkotnak. Ide tartoznak a nemzetséghez tartozó baktériumok Corynebacterium, Mycobacterium, Nocardia stb. A Nocardi-szerű aktinomicétákat az jellemzi, hogy a sejtfalban arabinóz, galaktóz, valamint mikolsav és nagy mennyiségű zsírsav található a sejtfalban. A mikolsavak és a sejtfallipidek határozzák meg a baktériumok savállóságát, különös tekintettel a mycobacteria tuberculosisra és a lepra (Ziehl-Neelsen szerint festve piros, a nem saválló baktériumok és szövetelemek, a köpet pedig kék színűek).
A kórokozó aktinomicéták aktinomikózist, nocardiát - nocardiózist, mycobacteriumot - tuberkulózist és leprát, korinebaktériumot - diftériát okoznak. A talajban elterjedtek az aktinomikéták szaprofita formái és a nocardiform actinomycetes, sok közülük antibiotikum termelő.
Mikoplazmák- kis baktériumok (0,15-1 µm), amelyeket csak egy szterolokat tartalmazó citoplazmatikus membrán vesz körül. Az osztályhoz tartoznak Mollicutes. A sejtfal hiánya miatt a mikoplazmák ozmotikusan érzékenyek. Különböző formájúak: coccoid, fonalas, lombik alakúak. Ezek a formák a mikoplazmák tiszta tenyészeteinek fáziskontrasztmikroszkópos képével láthatók. Sűrű táptalajon a mikoplazmák tükörtojásra emlékeztető kolóniákat alkotnak: egy központi, átlátszatlan részt a táptalajba merülve, és egy áttetsző kerületet kör alakban.
A mikoplazmák atipikus tüdőgyulladást okoznak emberben (Mycoplasma pneumoniae)és az urogenitális traktus elváltozásai
(M. hominis satöbbi.). A mikoplazmák nemcsak állatokban, hanem növényekben is betegségeket okoznak. A nem patogén képviselők is meglehetősen elterjedtek.
2.3. A gombák felépítése és osztályozása
A gomba a tartományhoz tartozik Eukarya, királyság Gombák (Mycota, Mycetes). A közelmúltban a gombákat és a protozoákat külön birodalmakra osztották: királyságra Eumycota(igazi gombák), királyság Chromistaés királyság Protozoa. Néhány korábban gombának vagy protozoonnak tartott mikroorganizmus egy új birodalomba került Chromista(króm lemezek). A gombák többsejtű vagy egysejtű, nem fotoszintetikus (klorofill-mentes) eukarióta mikroorganizmusok, vastag sejtfallal. Magjuk van nukleáris burokkal, citoplazma sejtszervekkel, citoplazmatikus membrán és többrétegű merev sejtfal, amely többféle poliszacharidból (mannánok, glükánok, cellulóz, kitin), valamint fehérjékből, lipidekből stb. áll. egy kapszula. A citoplazma membránja glikoproteineket, foszfolipideket és ergoszterolokat tartalmaz (szemben a koleszterinnel, az emlős szövetek fő szterinjével). A legtöbb gomba kötelező vagy fakultatív aerob.
A gombák elterjedtek a természetben, különösen a talajban. Egyes gombák hozzájárulnak a kenyér, sajt, tejsavtermékek és alkohol előállításához. Más gombák antimikrobiális antibiotikumokat (pl. penicillint) és immunszuppresszív gyógyszereket (pl. ciklosporint) termelnek. A gombákat genetikusok és molekuláris biológusok használják különféle folyamatok modellezésére. A fitopatogén gombák jelentős károkat okoznak a mezőgazdaságban, a gabonanövények és a gabonafélék gombás betegségeit okozzák. A gombák által okozott fertőzéseket mikózisoknak nevezik. Vannak hifális és élesztőgombák.
A hifális (penészes) gombák vagy hifomicéták 2-50 mikron vastag vékony szálakból, úgynevezett hifákból állnak, amelyek micéliummá vagy micéliummá (penészbe) fonódnak. A gomba testét tallusnak nevezik. Vannak demacium (pigmentált - barna vagy fekete) és hialin (nem pigmentált) hipomicéták. A tápanyagba benőtt hifák felelősek a gomba táplálásáért, ezeket vegetatív hifáknak nevezik. Hyphae, ra-
a szubsztrát felszíne felett állókat légi vagy szaporodási hifának nevezik (a szaporodásért felelősek). A telepek bolyhos megjelenésűek a légmicélium miatt.
Vannak alacsonyabb és magasabb gombák: a magasabb gombák hifáit válaszfalak, vagy lyukakkal ellátott válaszfalak választják el. Az alsóbbrendű gombák hifáinak nincs válaszfala, többmagvú sejtekről van szó, amelyeket cönocitikusnak neveznek (a görög szóból. koenos- egyetlen, közös).
Az élesztőgombákat (élesztőt) elsősorban 3-15 mikron átmérőjű egyedi ovális sejtek képviselik, telepeik a hifális gombákkal ellentétben kompakt megjelenésűek. Az ivaros szaporodás típusa szerint a magasabb gombák - ascomycetes és basidiomycetes - között oszlanak meg. Ivartalan szaporodáskor élesztő bimbózik vagy osztódik. Pseudohifákat és hamis micéliumot (pseudomycelium) alkothatnak megnyúlt sejtláncok - „kolbász” formájában. Az élesztőhöz hasonló, de ivaros szaporodási móddal nem rendelkező gombákat élesztőszerűnek nevezzük. Csak ivartalanul szaporodnak – bimbózás vagy hasadás útján. Az „élesztőszerű gombák” fogalmát gyakran azonosítják az „élesztő” fogalmával.
Sok gomba dimorfizmussal rendelkezik - a termesztési körülményektől függően képes hifális (micélium) vagy élesztőszerű növekedésre. A fertőzött szervezetben élesztőszerű sejtek (élesztőfázis) formájában szaporodnak, a táptalajokon hifákat és micéliumot képeznek. A dimorfizmus a hőmérsékleti tényezőhöz kapcsolódik: szobahőmérsékleten micélium képződik, és 37 ° C-on (emberi testhőmérsékleten) élesztőszerű sejtek képződnek.
A gombák ivarosan vagy ivartalanul szaporodnak. A gombák ivaros szaporodása ivarsejtek, ivaros spórák és más ivaros formák képződésével történik. A szexuális formákat teleomorfoknak nevezzük.
A gombák ivartalan szaporodása az anamorfoknak nevezett megfelelő formák képződésével történik. Az ilyen szaporodás bimbózás, a hifák és az ivartalan spórák feldarabolódása révén történik. Az endogén spórák (sporangiospórák) kerek szerkezetben - sporangiumban - érlelődnek. Az exogén spórák (konídiumok) a termő hifák, az úgynevezett konidiofórok csúcsain képződnek.
Sokféle konídium létezik. Az arthroconidiumok (arthrospórák), vagy talloconidiumok a hifák egyenletes elválasztásával és feldarabolásával, a blasztokonídiumok pedig a bimbózás eredményeként jönnek létre. A kis egysejtű konídiumokat mikrokonídiumoknak, a nagy többsejtűeket makrokonídiumoknak nevezzük. A gombák ivartalan formái közé tartoznak a chlamydoconidia vagy a chlamydospórák (vastag falú nagy nyugvósejtek vagy kis sejtek komplexuma) is.
Vannak tökéletes és tökéletlen gombák. A tökéletes gombák szexuális szaporodási módszerrel rendelkeznek; ezek közé tartoznak a zygomyceták (Zygomycota), ascomycetes (Ascomycota)és bazidiomycetes (Basidiomycota). A tökéletlen gombák csak ivartalan szaporodással rendelkeznek; Ide tartozik a formális hagyományos gombatípus/-csoport - deuteromycetes (Deiteromycota).
A zygomycetes az alsóbb gombákhoz (nem szeptummicélium) tartozik. Közéjük tartoznak a nemzetségek képviselői Mucor, Rhizopus, Rhizomucor, Absidia, Basidiobolus, Conidiobolus. Talajban és levegőben oszlik el. A tüdő, az agy és más emberi szervek zygomycosisát (mukoromikózisát) okozhatják.
A zygomyceták ivartalan szaporodása során a termőhifákon (sporangioforokon) sporangium képződik - gömb alakú megvastagodás, számos sporangiospórát tartalmazó héjjal (2.6., 2.7. ábra). Az ivaros szaporodás a zygomycetákban zigospórák segítségével történik.
Az Ascomycetes (erszényes gombák) septate micéliummal rendelkeznek (kivéve az egysejtű élesztőgombákat). Nevüket a fő termőszervről kapták - a bursáról vagy aszkuszról, amely 4 vagy 8 haploid ivaros spórát (aszkospórát) tartalmaz.
Az Ascomycetes a nemzetség egyes képviselőit (teleomorfokat) foglalja magában AspergillusÉs Penicillium. A legtöbb gomba nemzetség Aspergillus, Penicillium anamorfok, azaz. csak tehetetlenül szaporodnak
 Rizs. 2.6. A nemzetséghez tartozó gombák Mucor(a kép: A.S. Bykov)
Rizs. 2.6. A nemzetséghez tartozó gombák Mucor(a kép: A.S. Bykov)
 Rizs. 2.7. A nemzetséghez tartozó gombák Rhizopus. Sporangiumok, sporangiospórák és rizoidok fejlődése
Rizs. 2.7. A nemzetséghez tartozó gombák Rhizopus. Sporangiumok, sporangiospórák és rizoidok fejlődése
az első módon ivartalan spórák - konídiumok (2.8. ábra, 2.9. ábra) segítségével, és e tulajdonság alapján a tökéletlen gombák közé kell sorolni. A nemzetséghez tartozó gombákban Aspergillus a termő hifák, konidioforok végein megvastagodások - sterigmák, phialidák vannak, amelyeken konídiumláncok képződnek („vízpenész”).
A nemzetséghez tartozó gombákban Penicillium(bojt) a termő hifa egy ecsettel hasonlít, mivel belőle (a konidioforon) megvastagodások képződnek, amelyek kisebb struktúrákra - sterigmákra, phialidákra - ágaznak, amelyeken konídiumláncok vannak. Az Aspergillus egyes fajai aspergillózist és aflatoxikózist, a Penicillium pedig penicilliózist okozhat.
Az ascomycetes képviselői a nemzetség teleomorfjai Trichophyton, Microsporum, Histoplasma, Blastomyces, valamint a remegés
 Rizs. 2.8. A nemzetséghez tartozó gombák Penicillium. A fialidokból konídiumláncok nyúlnak ki
Rizs. 2.8. A nemzetséghez tartozó gombák Penicillium. A fialidokból konídiumláncok nyúlnak ki
 Rizs. 2.9. A nemzetséghez tartozó gombák Aspergillus fumigatus. A fialidokból konídiumláncok nyúlnak ki
Rizs. 2.9. A nemzetséghez tartozó gombák Aspergillus fumigatus. A fialidokból konídiumláncok nyúlnak ki
A Basidiomycetes közé tartoznak a kalapgombák. Szeptált micéliummal rendelkeznek, és a bazídiumról leválva ivaros spórákat képeznek - bazidiospórák - a micélium terminális sejtje, amely homológ az ascusszal. A bazidiomicéták közé tartoznak néhány élesztőgomba, például a teleomorfok Cryptococcus neoformans.
A Deuteromycetes tökéletlen gombák (Fungi imperfecti, anamorf gombák, konídium gombák). Ez a gombák feltételes, formális taxonja, egyesíti azokat a gombákat, amelyek nem rendelkeznek ivaros szaporodással. A közelmúltban a „deuteromycetes” kifejezés helyett a „mitospórusos gombák” kifejezést javasolták - olyan gombák, amelyek nem ivaros spórákkal szaporodnak, pl. mitózis által. Amikor megállapítják a tökéletlen gombák ivaros szaporodásának tényét, átkerülnek az ismert típusok egyikébe - Ascomycota vagy Basidiomycota a név hozzárendelése egy teleomorf alakhoz. A deuteromyceták septatikus micéliummal rendelkeznek, és csak a konídiumok ivartalan képződésével szaporodnak. A deuteromyceták közé tartoznak a tökéletlen élesztőgombák (élesztőszerű gombák), például a nemzetség egyes gombái Candida, a bőrt, a nyálkahártyákat és a belső szerveket érinti (candidiasis). Ovális alakúak, 2-5 mikron átmérőjűek, bimbózással osztódnak, megnyúlt sejtláncok formájában pszeudohifákat (pseudomycelium) alkotnak, és néha hifákat alkotnak. Mert Candida albicans jellemző a chlamydospórák képződése (2.10. ábra). A deuteromyceteshez tartoznak a nem ivaros szaporodási móddal nem rendelkező gombák is, amelyek a nemzetségbe tartoznak. Epidermophyton, Coccidioides, Paracoccidioides, Sporothrix, Aspergillus, Phialophora, Fonsecaea, Exophiala, Cladophialophora, Bipolaris, Exerohilum, Wangiella, Alrernaria satöbbi.
 Rizs. 2.10. A nemzetséghez tartozó gombák Candida albicans(a kép: A.S. Bykov)
Rizs. 2.10. A nemzetséghez tartozó gombák Candida albicans(a kép: A.S. Bykov)
2.4. A protozoonok szerkezete és osztályozása
A legegyszerűbbek a tartományhoz tartoznak Eukarya,állatvilág (Állat), alkirályság Protozoa. A közelmúltban javasolták a protozoonok királyi rangba való besorolását Protozoa.
A protozoon sejtet membrán (pellikulum) veszi körül - az állati sejtek citoplazmatikus membránjának analógja. Van egy magja nukleáris burokkal és sejtmaggal, egy citoplazma, amely tartalmazza az endoplazmatikus retikulumot, mitokondriumokat, lizoszómákat és riboszómákat. A protozoonok mérete 2 és 100 mikron között van. Romanovsky-Giemsa szerint festve a protozoon magja vörös, a citoplazma kék. A protozoonok flagellák, csillók vagy pszeudopodiák segítségével mozognak, némelyikük emésztő- és összehúzó (kiválasztó) vakuólumokkal rendelkezik. Fagocitózis vagy speciális struktúrák kialakulása következtében táplálkozhatnak. A táplálkozás típusa alapján heterotrófokra és autotrófokra osztják őket. Számos protozoa (dizentériás amőba, Giardia, Trichomonas, Leishmania, Balantidia) nőhet natív fehérjéket és aminosavakat tartalmazó táptalajokon. Tenyésztésükhöz sejtkultúrákat, csirkeembriókat és laboratóriumi állatokat is használnak.
A protozoonok ivartalanul szaporodnak - kettős vagy többszörös osztódással (szkizogónia), néhány pedig ivarosan is (sporogónia). Egyes protozoonok extracellulárisan (Giardia), mások intracellulárisan (Plasmodium, Toxoplasma, Leishmania) szaporodnak. A protozoonok életciklusát szakaszok jellemzik - a trophozoit szakasz és a ciszta szakasz kialakulása. A ciszták alvó stádiumok, ellenállnak a hőmérséklet és a páratartalom változásainak. A ciszták savállóak Sarcocystis, CryptosporidiumÉs Isospora.
Korábban az emberben betegségeket okozó protozoonokat 4 típus képviselte 1 ( Sarcomastigophora, Apicomplexa, Ciliophora, Microspora). Ezeket a típusokat a közelmúltban nagyobb számba sorolták át, és új királyságok alakultak ki - ProtozoaÉs Chromista(2.2. táblázat). Egy új királyság felé Chromista(chromovics) tartalmazott néhány protozoont és gombát (blasztociszták, oomyceták és Rhinosporidium seeberi). Királyság Protozoa magában foglalja az amőbákat, flagellátákat, sporozoákat és csillós állatokat. Különböző típusokra oszthatók, köztük amőbák, flagellátok, sporozoák és csillósok.
2.2. táblázat. A királyságok képviselői ProtozoaÉs Chromista, orvosi jelentőségű
 1 Típus Sarcomastigophora altípusokból állt SarcodinaÉs Mastigophora. Altípus Sarcodina(sarcodaceae) magában foglalja a vérhas amőbát és az altípust Mastigophora(flagellates) - tripanoszómák, leishmania, lamblia és trichomonas. típus Apicomplexa osztályba foglalt Sporozoa(spórák), amelyek magukban foglalták a malária plazmódiáit, a toxoplazmát, a cryptosporidiumot stb. Ciliophora magában foglalja a balantidia és a típust Microspora- microsporidia.
1 Típus Sarcomastigophora altípusokból állt SarcodinaÉs Mastigophora. Altípus Sarcodina(sarcodaceae) magában foglalja a vérhas amőbát és az altípust Mastigophora(flagellates) - tripanoszómák, leishmania, lamblia és trichomonas. típus Apicomplexa osztályba foglalt Sporozoa(spórák), amelyek magukban foglalták a malária plazmódiáit, a toxoplazmát, a cryptosporidiumot stb. Ciliophora magában foglalja a balantidia és a típust Microspora- microsporidia.
A táblázat vége. 2.2
 Az amőbák közé tartozik az emberi amőbiázis - amőbás vérhas - kórokozója (Entamoeba histolytica), szabadon élő és nem patogén amőbák (bélamőba stb.). Az amőbák ivartalanul binárisan szaporodnak. Életciklusuk egy trofozoit állapotból (növekvő, mozgékony sejt, gyengén stabil) és egy ciszta szakaszból áll. A trophozoiták pszeudopodiák segítségével mozognak, amelyek felfogják és a tápanyagokat a citoplazmába merítik. Tól től
Az amőbák közé tartozik az emberi amőbiázis - amőbás vérhas - kórokozója (Entamoeba histolytica), szabadon élő és nem patogén amőbák (bélamőba stb.). Az amőbák ivartalanul binárisan szaporodnak. Életciklusuk egy trofozoit állapotból (növekvő, mozgékony sejt, gyengén stabil) és egy ciszta szakaszból áll. A trophozoiták pszeudopodiák segítségével mozognak, amelyek felfogják és a tápanyagokat a citoplazmába merítik. Tól től
A trofozoit cisztát képez, amely ellenáll a külső tényezőknek. A bélbe kerülve trophozoitává alakul.
A flagellákra jellemző a flagellák jelenléte: a Leishmania-ban egy flagellum, a Trichomonas-ban 4 szabad flagella és egy rövid, hullámos membránhoz kapcsolódó flagellum található. Ők:
A vér és a szövetek lobogói (Leishmania - a leishmaniasis kórokozói; Trypanosomes - az alvási betegség és a Chagas-kór kórokozói);
Bélkorongok (giardia - a giardiasis kórokozója);
Az urogenitális traktus zászlói (Trichomonas vaginalis - a trichomoniasis kórokozója).
A csillósokat a balantidiumok képviselik, amelyek az emberi vastagbelet érintik (balantidiasis dizentéria). A balantidia trofozoit és ciszta stádiumú. A trofozoit mozgékony, számos csillóval rendelkezik, vékonyabb és rövidebb, mint a flagellák.
2.5. A vírusok szerkezete és osztályozása
A vírusok a királysághoz tartozó legkisebb mikrobák Virae(a lat. vírus- I). Nem rendelkeznek sejtszerkezettel és állnak
A vírusok szerkezetét kis méretük miatt mind a virionok, mind azok ultravékony metszete elektronmikroszkópos vizsgálatával vizsgálják. A vírusok (virionok) méretét közvetlenül elektronmikroszkóppal vagy közvetetten, ismert pórusátmérőjű szűrőkön végzett ultraszűréssel vagy ultracentrifugálással határozzák meg. A vírusok mérete 15-400 nm (1 nm egyenlő 1/1000 μm): a kis vírusok, amelyek mérete hasonló a riboszómák méretéhez, a parvovírusok és a poliovírusok, a legnagyobbak pedig a variola vírusok (350) nm). A vírusok virionjaik alakjában különböznek, amelyek pálcikák (dohánymozaik vírus), golyók (veszettség vírus), gömbök (poliomyelitis vírusok, HIV), fonalak (filovírusok), spermiumok (sok bakteriofág) formájúak.
A vírusok szerkezetük és tulajdonságaik sokféleségével ámulatba ejtik a képzeletet. Ellentétben a sejtgenomokkal, amelyek egységes kétszálú DNS-t tartalmaznak, a vírusgenomok rendkívül változatosak. Vannak olyan DNS és RNS vírusok, amelyek haploidok, azaz. egy génkészlettel rendelkezik. Csak a retrovírusok rendelkeznek diploid genommal. A vírusok genomja 6-200 gént tartalmaz, és különféle típusú nukleinsavak képviselik: kétszálú, egyszálú, lineáris, körkörös, fragmentált.
Az egyszálú RNS-vírusok között különbséget tesznek a genomiális plusz szálú RNS és a mínusz szálú RNS között (RNS polaritás). Ezen vírusok RNS-ének plusz szála (pozitív szála) a genomiális (örökletes) funkción kívül információs vagy hírvivő RNS (mRNS, vagy mRNS) funkciót is ellát; ez a fehérjeszintézis mátrixa a fertőzött sejt riboszómáin. A plusz szálú RNS fertőző: érzékeny sejtekbe kerülve fertőző folyamatot idézhet elő.
cess. Az RNS-vírusok mínusz szála (negatív szála) csak örökletes funkciót lát el; A fehérjeszintézishez egy komplementer szálat szintetizálnak az RNS mínusz szálán. Egyes vírusok ambipoláris RNS genommal rendelkeznek (ambisense görögből ambi- mindkét oldalon kettős komplementaritás), azaz. plusz és mínusz RNS szegmenseket tartalmaz.
Vannak egyszerű vírusok (például hepatitis A vírus) és összetett vírusok (például influenzavírusok, herpesz, koronavírusok).
Az egyszerű vagy nem burkolt vírusok csak nukleinsavat tartalmaznak, amely a kapszidnak nevezett fehérjeszerkezethez kapcsolódik (a latin. capsa- eset). A nukleinsavhoz kapcsolódó fehérjéket nukleoproteineknek nevezik, a víruskapszidfehérjéknek a virális nukleinsavval való asszociációját pedig nukleokapszidnak nevezik. Néhány egyszerű vírus kristályokat képezhet (pl. ragadós száj- és körömfájás vírus).
A kapszid ismétlődő morfológiai alegységeket - kapszomereket - tartalmaz, amelyek több polipeptidből állnak. A virion nukleinsavja a kapszidhoz kötve nukleokapszidot képez. A kapszid megvédi a nukleinsavat a lebomlástól. Az egyszerű vírusokban a kapszid részt vesz a gazdasejthez való kapcsolódásban (adszorpcióban). Az egyszerű vírusok pusztulása (lízise) következtében elhagyják a sejtet.
A komplex, vagy burkolt vírusok (2.11. ábra) a kapszidon kívül membrán kettős lipoprotein burokkal (szinonimája: szuperkapszid vagy peplos) is rendelkeznek, amelyet a virion sejtmembránon keresztül történő bimbózása révén szereznek meg, például a sejtmembránon keresztül. plazmamembrán, magmembrán vagy endoplazmatikus retikulum membrán. A vírusburok glikoprotein-csúcsokat tartalmaz,
vagy tüskék, peplomerek. A héj éterrel és más oldószerekkel történő megsemmisítése inaktiválja a komplex vírusokat. Egyes vírusok héja alatt mátrixfehérje (M protein) található.
A virionok spirális, ikozaéderes (köbös) vagy összetett típusú kapszid (nukleokapszid) szimmetriával rendelkeznek. A spirális típusú szimmetria a nukleokapszid spirális szerkezetéből adódik (például influenzavírusoknál, koronavírusoknál): a kapszomerek a nukleinsavval együtt spirálisan helyezkednek el. Az ikozaéder típusú szimmetria annak köszönhető, hogy a vírus nukleinsavat tartalmazó kapszidból izometrikusan üreges test képződik (például a herpeszvírusban).
A kapszid és a héj (szuperkapszid) megvédi a virionokat a környezeti hatásoktól, és meghatározza receptorfehérjeik szelektív kölcsönhatását (adszorpcióját) bizonyos
 Rizs. 2.11. Az ikozaéderes (a) és spirális (b) kapszidú burokkal rendelkező vírusok szerkezete
Rizs. 2.11. Az ikozaéderes (a) és spirális (b) kapszidú burokkal rendelkező vírusok szerkezete
sejteket, valamint a virionok antigén és immunogén tulajdonságait.
A vírusok belső szerkezetét magnak nevezzük. Az adenovírusokban a mag a DNS-hez kapcsolódó hisztonszerű fehérjékből, a reovírusoknál a belső kapszid fehérjéiből áll.
A Nobel-díjas D. Baltimore javasolta az mRNS-szintézis mechanizmusán alapuló baltimore-i osztályozási rendszert. Ez a besorolás a vírusokat 7 csoportba sorolja (2.3. táblázat). Vírusok Taxonómiájának Nemzetközi Bizottsága (ICTV) egy univerzális osztályozási rendszert fogadott el, amely olyan taxonómiai kategóriákat használ, mint például a család (a név végződése viridae), alcsalád (a név számra végződik virinae), nemzetség (a név számra végződik vírus). A vírusfajták nem kaptak binomiális nevet, mint a baktériumok. A vírusokat a nukleinsav típusa (DNS vagy RNS), szerkezete és a szálak száma alapján osztályozzák. Kétszálú vagy egyszálú nukleinsavak vannak; pozitív (+), negatív (-) nukleinsav polaritás vagy vegyes nukleinsav polaritás, ambipoláris (+, -); lineáris vagy cirkuláris nukleinsav; fragmentált vagy nem fragmentált nukleinsav. Figyelembe veszik a virionok méretét és morfológiáját, a kapszomerek számát és a nukleokapszid szimmetriájának típusát, a burok (szuperkapszid) jelenlétét, az éterrel és dezoxikoláttal szembeni érzékenységet, a szaporodási helyet a sejtben, az antigén tulajdonságokat stb. figyelembe.
2.3. táblázat. Főbb orvosi jelentőségű vírusok
 A táblázat folytatása. 2.3
A táblázat folytatása. 2.3
 A táblázat vége. 2.3
A táblázat vége. 2.3
 A vírusok állatokat, baktériumokat, gombákat és növényeket fertőznek meg. Az emberi fertőző betegségek fő kórokozóiként a vírusok a karcinogenezis folyamataiban is részt vesznek, és többféle módon terjedhetnek, többek között a méhlepényen keresztül (rubeola vírus, citomegalovírus stb.), az emberi magzatot érintve. Ezek fertőzés utáni szövődményekhez is vezethetnek - szívizomgyulladás, hasnyálmirigy-gyulladás, immunhiány stb.
A vírusok állatokat, baktériumokat, gombákat és növényeket fertőznek meg. Az emberi fertőző betegségek fő kórokozóiként a vírusok a karcinogenezis folyamataiban is részt vesznek, és többféle módon terjedhetnek, többek között a méhlepényen keresztül (rubeola vírus, citomegalovírus stb.), az emberi magzatot érintve. Ezek fertőzés utáni szövődményekhez is vezethetnek - szívizomgyulladás, hasnyálmirigy-gyulladás, immunhiány stb.
A vírusokon kívül a nem sejtes életformák közé tartoznak a prionok és a viroidok. A viroidok cirkuláris, szupertekervényes RNS kis molekulái, amelyek nem tartalmaznak fehérjét és növényi betegségeket okoznak. A kóros prionok olyan fertőző fehérjerészecskék, amelyek a normál sejt prionfehérje szerkezetében bekövetkező változások következtében speciális konformációs betegségeket okoznak. PrP c), amely jelen van az állatok és az emberek szervezetében. PrP-vel szabályozó funkciókat lát el. A normál priongén (PrP gén) kódolja, amely az emberi 20-as kromoszóma rövid karján található. A prionbetegségek fertőző szivacsos agyvelőbántalmakként fordulnak elő (Creutzfeldt-Jakob-kór, kuru stb.). Ebben az esetben a prionfehérje egy másik, fertőző formát kap, amelyet a következőképpen jelölünk PrP sc(sc from surlókór- a surlókór a juhok és kecskék prionfertőzése). Ez a fertőző prionfehérje rostszerű megjelenésű, és harmadlagos vagy kvaterner szerkezetében különbözik a normál prionfehérjétől.
Önfelkészítő feladatok (önkontroll)
A. Jelölje meg azokat a mikrobákat, amelyek prokarióták:
2. Vírusok.
3. Baktériumok.
4. Prionok.
B. Vegye figyelembe a prokarióta sejt megkülönböztető jellemzőit:
1. 70S riboszómák.
2. A peptidoglikán jelenléte a sejtfalban.
3. Mitokondriumok jelenléte.
4. Diploid génkészlet.
BAN BEN. Jelölje meg a peptidoglikán összetevőit:
1. Teichoinsavak.
2. N-acetil-glükózamin.
3. Lipopolizarid.
4. Tetrapeptid.
G. Vegye figyelembe a gram-negatív baktériumok sejtfalának szerkezeti jellemzőit:
1. Mezodiaminopimelinsav.
2. Teichoinsavak.
4. Porin fehérjék.
D. Nevezze meg a spórák funkcióit a baktériumokban:
1. A faj megőrzése.
2. Hőállóság.
3. Az aljzat diszpergálása.
4. Szaporodás.
1. Rickettsia.
2. Actinomycetes.
3. Spirochetes.
4. Chlamydia.
ÉS. Nevezze meg az aktinomyceták jellemzőit:
1. Termolabilis spóráik vannak.
2. Gram-pozitív baktériumok.
3. Nincs sejtfal.
4. Tekervényes alakjuk van.
Z. Nevezze meg a spirocheták jellemzőit:
1. Gram-negatív baktériumok.
2. Motoros fibrilláris berendezéssel rendelkeznek.
3. Tekervényes alakjuk van.
ÉS. Nevezze meg azokat a protozoonokat, amelyek apikális komplexummal rendelkeznek, amely lehetővé teszi számukra, hogy behatoljanak a sejtbe:
1. Malária plazmódium.
3. Toxoplazma.
4. Cryptosporidium.
NAK NEK. Nevezze meg a komplex vírusok megkülönböztető jellemzőit:
1. Kétféle nukleinsav.
2. Lipid membrán jelenléte.
3. Dupla kapszid.
4. Nem strukturális fehérjék jelenléte. L. Jelölje meg a magasabb gombákat:
1. Mucor.
2. Candida.
3. Penicillium.
4. Aspergillus.
A baktériumsejtnek nincs sejtmagja, a kromoszómák szabadon helyezkednek el a citoplazmában. Ezenkívül a baktériumsejtből hiányoznak a membránszervecskék: mitokondriumok, EPS, Golgi-készülék stb. A sejtmembrán külső részét sejtfal borítja.



A legtöbb baktérium passzívan mozog, víz- vagy légáramlatok segítségével. Csak néhányuknak van mozgásszervezete - flagella. A prokarióta flagellák nagyon egyszerű szerkezetűek, és flagellin fehérjéből állnak, amely 10-20 nm átmérőjű üreges hengert alkot. Becsavarják a táptalajt, előrehajtva a sejtet. Úgy tűnik, ez az egyetlen olyan szerkezet a természetben, amely a kerék elvét használja.
Alakjuk alapján a baktériumokat több csoportra osztják:
Cocci (kerek alakú);
- bacillusok (rúd alakúak);
- spirilla (spirál alakú);
- vibrios (vessző alakú).
A légzés módja szerint a baktériumokat aerobokra (a legtöbb baktérium) és anaerobokra (tetanusz, botulizmus, gázgangréna kórokozói) osztják. Az előbbieknek oxigénre van szükségük a légzéshez, az oxigén haszontalan vagy akár mérgező is.

A prokarióta sejt felépítése. A prokarióta sejt sokkal egyszerűbb, mint az állati és növényi sejtek. Kívülről sejtfal borítja, amely védő, formáló és szállító funkciókat lát el. A sejtfal merevségét a murein biztosítja. Néha a baktériumsejt tetején kapszula vagy nyálkahártya borítja.
A baktériumok protoplazmáját az eukariótákhoz hasonlóan plazmamembrán veszi körül. A membrán zsákszerű, tubuláris vagy lamellás invaginációi a légzési folyamatban részt vevő mezoszómákat, bakterioklorofillt és más pigmenteket tartalmaznak.
A prokarióták genetikai anyaga nem alkot magot, hanem közvetlenül a citoplazmában található. A bakteriális DNS egyetlen kör alakú molekula, amelyek mindegyike több ezer és millió nukleotidpárból áll. A baktériumsejt genomja sokkal egyszerűbb, mint a fejlettebb lények sejtjeié: átlagosan több ezer gént tartalmaz a bakteriális DNS.
A prokarióta sejtekben nincs endoplazmatikus retikulum, és a riboszómák szabadon lebegnek a citoplazmában. A prokariótáknak nincs mitokondriumuk; Feladatukat részben a sejtmembrán látja el.
A baktériumok mobilitását a flagellák biztosítják. A baktériumok körülbelül 20 percenkénti osztódással szaporodnak (kedvező körülmények között). A DNS-t replikálják, és minden leánysejt megkapja a saját DNS-másolatát. Nem osztódó sejtek között is lehetséges DNS átvitel ("csupasz" DNS befogásával, bakteriofágok használatával, vagy konjugációval, amikor a baktériumok kopulációs fimbriákkal kapcsolódnak egymáshoz), de ez nem növeli az egyedek számát. A szaporodást a napsugárzás és saját létfontosságú tevékenységük termékei akadályozzák.
A baktériumok viselkedése nem különösebben bonyolult. A kémiai receptorok rögzítik a környezet savasságának és a különféle anyagok koncentrációjának változásait: cukrok, aminosavak, oxigén. Sok baktérium reagál a hőmérséklet vagy a fény változására, és néhány baktérium érzékeli a Föld mágneses terét. Kedvezőtlen körülmények között a baktériumot sűrű héj borítja, a citoplazma kiszárad, élettevékenysége szinte megszűnik. Ebben az állapotban a baktériumspórák órákig mély vákuumban maradhatnak, és elviselik a –240 °C és +100 °C közötti hőmérsékletet.