Et mitte ainult mõista raku struktuuriliste tunnuste olulisust, vaid, mis kõige tähtsam, mõista selle üksikute komponentide ja kogu raku funktsionaalseid funktsioone, et ühendada raku morfoloogia uurimine selle struktuuri ja töö peamised biokeemilised ja geneetilised tunnused, et uurida rakku täpselt kaasaegse rakubioloogia positsioonidega, on vaja vähemalt põgusalt meelde tuletada peamised molekulaarbioloogilised mustrid, veel kord lühidalt viidata keskse dogma sisule. molekulaarbioloogia.
Rakk kui selline täidab palju erinevaid funktsioone. Nagu me juba ütlesime, on mõned neist üldised rakulised, mõned on erilised, iseloomulikud spetsiaalsetele rakutüüpidele. Nende funktsioonide peamised töömehhanismid on valgud või nende kompleksid teiste bioloogiliste makromolekulidega, nagu nukleiinhapped, lipiidid ja polüsahhariidid. Näiteks on teada, et erinevate ainete, ioonidest makromolekulideni, transpordiprotsessid rakus on määratud spetsiaalsete valkude või lipoproteiinikomplekside tööga, mis on osa plasmast ja muudest. rakumembraanid. Peaaegu kõik erinevate valkude sünteesi, lagunemise, ümberkorraldamise protsessid, nukleiinhapped, lipiidid, süsivesikud tekivad iga üksiku reaktsiooni jaoks spetsiifiliste valkude-ensüümide aktiivsuse tulemusena. Üksikute bioloogiliste monomeeride, nukleotiidide, aminohapete süntees, rasvhapped, suhkruid ja muid ühendeid teostab ka tohutu hulk spetsiifilisi ensüüme – valke. Kontraktsiooni, mis viib rakkude liikuvuseni või ainete ja struktuuride liikumiseni rakkude sees, teostavad ka spetsiaalsed kontraktiilsed valgud. Paljud raku reaktsioonid vastuseks välisteguritele (viirused, hormoonid, võõrvalgud jne) saavad alguse nende tegurite koostoimest spetsiaalsete raku retseptorvalkudega.
Valgud on peaaegu kõigi rakustruktuuride põhikomponendid. Trobikond keemilised reaktsioonid raku sees määravad paljud ensüümid, millest igaüks viib ühe või mitu eraldi reaktsiooni. Iga üksiku valgu struktuur on rangelt spetsiifiline, mis väljendub nende esmase struktuuri spetsiifilisuses - aminohapete järjestuses piki polüpeptiidi, valguahelat. Veelgi enam, selle aminohappejärjestuse spetsiifilisus kordub eksimatult kõigis selle rakuvalgu molekulides.
Sellise korrektsuse valguahelas üheselt mõistetava aminohappejärjestuse reprodutseerimisel määrab selle geenipiirkonna DNA struktuur, mis lõppkokkuvõttes vastutab selle valgu struktuuri ja sünteesi eest. Need ideed on molekulaarbioloogia peamine postulaat, selle "dogma". Teave tulevase valgu molekuli kohta edastatakse selle sünteesikohtadesse (ribosoomidesse) vahendaja - messenger RNA (mRNA) kaudu, mille nukleotiidide koostis peegeldab DNA geenipiirkonna koostist ja nukleotiidjärjestust. Ribosoomi on ehitatud polüpeptiidahel, mille aminohapete järjestuse määrab mRNA nukleotiidide järjestus, nende kolmikute järjestus. Seega rõhutab molekulaarbioloogia keskne dogma teabe ühesuunalist edastamist: ainult DNA-st valku vaheühendi - mRNA (DNA → mRNA → valk) abil. Mõnede RNA-d sisaldavate viiruste puhul võib infoedastusahel järgida skeemi RNA → mRNA → valk. Asja olemust see ei muuda, kuna ka siin on määravaks lüliks nukleiinhape. Valgust nukleiinhappeks, DNA-ks või RNA-ks määramise vastupidised teed ei ole teada.
Et jätkata valgusünteesi kõigi etappidega seotud rakustruktuuride uurimist, peame põgusalt peatuma peamistel protsessidel ja komponentidel, mis seda nähtust määravad.
Praegu põhineb kaasaegsed ideed valkude biosünteesi kohta saame anda selle keerulise ja mitmeetapilise protsessi üldise skemaatilise diagrammi (joonis 16).
Peamine, "käsu" roll valkude spetsiifilise struktuuri määramisel kuulub desoksüribonukleiinhappele - DNA-le. DNA molekul on ülipikk lineaarne struktuur, mis koosneb kahest omavahel põimunud polümeeri ahelast. Nende ahelate koostisosad – monomeerid – on nelja tüüpi desoksüribonukleotiidid, mille vaheldumine või järjestus piki ahelat on ainulaadne ja spetsiifiline iga DNA molekuli ja selle iga sektsiooni jaoks. Erinevate valkude sünteesi eest vastutavad erinevad piisavalt pikad DNA molekuli lõigud. Seega saab sünteesi määrata üks DNA molekul suur hulk funktsionaalselt ja keemiliselt erinevad rakuvalgud. Igat tüüpi valgu sünteesi eest vastutab ainult teatud osa DNA molekulist. Sellist DNA molekuli piirkonda, mis on seotud ühe konkreetse valgu sünteesiga rakus, nimetatakse sageli terminiga "tsistron". Praegu peetakse tsistroni mõistet samaväärseks geeni mõistega. Geeni ainulaadne struktuur - selle nukleotiidide teatud järjestikuses paigutuses piki ahelat - sisaldab kogu teavet ühe vastava valgu struktuuri kohta.
Alates üldine skeem valgusüntees, on näha (vt joonis 16), et lähtepunkt, millest algab infovoog valkude biosünteesiks rakus, on DNA. Järelikult on DNA see, mis sisaldab esmast teavet, mida tuleb säilitada ja reprodutseerida rakust rakku, põlvest põlve.
Lühidalt ladustamise probleemi käsitlemine geneetiline teave, st. DNA lokaliseerimise kohta rakus võime öelda järgmist. Juba ammu on teada, et erinevalt kõigist teistest valke sünteesiva aparaadi komponentidest on DNA-l eriline, väga piiratud lokalisatsioon: selle asukohaks kõrgemate (eukarüootsete) organismide rakkudes saab olema raku tuum. Madalamates (prokarüootsetes) organismides, millel ei ole hästi moodustunud rakutuum, segatakse DNA ka ülejäänud protoplasmast ühe või mitme kompaktse nukleotiidmoodustise kujul. Sellega täielikult kooskõlas on eukarüootide tuuma või prokarüootide nukleoidi peetud pikka aega geenide mahutiks, ainulaadseks rakuliseks organelliks, mis kontrollib organismide pärilike tunnuste rakendamist ja nende edasikandumist põlvkondade kaupa.
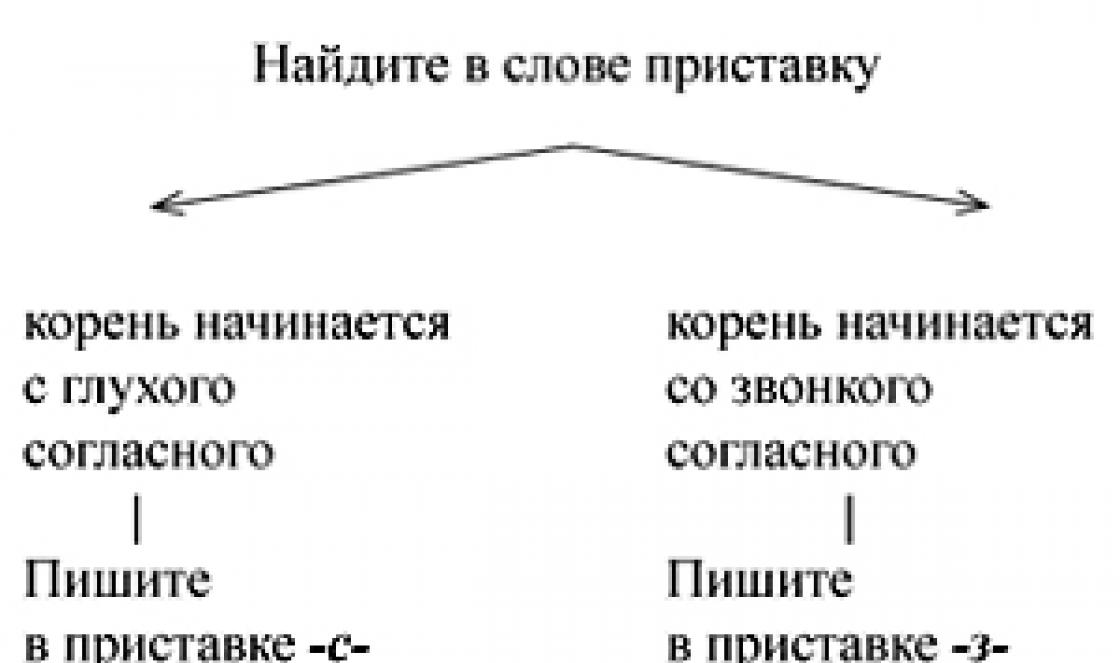
DNA makromolekulaarse struktuuri peamine põhimõte on nn komplementaarsuse printsiip (joonis 17). Nagu juba mainitud, koosneb DNA molekul kahest omavahel põimunud ahelast. Need ahelad on üksteisega seotud nende vastandlike nukleotiidide interaktsiooni kaudu. Samas on struktuursetel põhjustel sellise kaheahelalise struktuuri olemasolu võimalik vaid juhul, kui mõlema ahela vastassuunalised nukleotiidid on steeriliselt komplementaarsed, s.t. saab olema nende ruumiline struktuur täiendavad üksteist. Sellised komplementaarsed - komplementaarsed - nukleotiidide paarid on paar A-T(adeniin-tüümiin) ja paar G-C (guaniin-tsütosiin).
Seetõttu on selle komplementaarsuse põhimõtte kohaselt, kui DNA molekuli ühes ahelas on teatud nelja tüüpi nukleotiidide järjestus, siis teises ahelas määratakse nukleotiidide järjestus üheselt, nii et esimese ahela iga A. vastab T-le teises ahelas, igale T-le esimeses ahelas - A teises ahelas, igale G esimeses ahelas - C-le teises ahelas ja igale C-le esimeses ahelas - G-le teises ahelas.
See DNA molekuli kaheahelalise struktuuri aluseks olev struktuurne põhimõte võimaldab hõlpsasti mõista algse struktuuri täpset reprodutseerimist, s.o. molekuli ahelatesse salvestatud teabe täpne reprodutseerimine nelja tüüpi nukleotiidide spetsiifilise järjestuse kujul. Tõepoolest, uute DNA molekulide süntees rakus toimub ainult olemasolevate DNA molekulide baasil. Sel juhul hakkavad algse DNA molekuli kaks ahelat ühest otsast lahknema ja igal eraldatud üheahelalisel lõigul hakkab teine ahel söötmes sisalduvatest vabadest nukleotiididest kogunema rangelt kooskõlas täiendavuse põhimõtet. Algse DNA molekuli kahe ahela lahknemise protsess jätkub ja vastavalt sellele täiendatakse mõlemat ahelat komplementaarsete ahelatega. Selle tulemusena (nagu on näha joonisel 17) ilmub ühe DNA molekuli asemel kaks, mis on täpselt identsed originaaliga. Igas saadud "tütar" DNA molekulis on üks ahel täielikult tuletatud originaalist ja teine on äsja sünteesitud.
Tuleb rõhutada, et täpse paljunemise potentsiaal on omane DNA kaheahelalisele komplementaarsele struktuurile kui sellisele ja selle avastamine on loomulikult bioloogia üks peamisi saavutusi.
Kuid DNA reprodutseerimise (reduplikatsiooni) probleem ei piirdu väitega selle struktuuri võimaliku võime kohta oma struktuuri täpselt reprodutseerida. nukleotiidjärjestus. Fakt on see, et DNA ise ei ole üldse isepaljunev molekul. Sünteesiprotsessi läbiviimiseks - DNA reprodutseerimiseks vastavalt ülalkirjeldatud skeemile - on vajalik spetsiaalse ensümaatilise kompleksi, mida nimetatakse DNA polümeraasiks, aktiivsus. Just see ensüüm liigub järjestikku DNA molekuli ühest otsast teise, kahe ahela eraldamise protsess koos nende vabade nukleotiidide samaaegse polümerisatsiooniga vastavalt komplementaarsele põhimõttele. Seega määrab DNA sarnaselt maatriksiga ainult nukleotiidide järjestuse sünteesitud ahelates ja protsessi ise viib läbi valk. Ensüümi töö DNA replikatsiooni ajal on praegu üks huvitavamaid probleeme. On tõenäoline, et DNA polümeraas roomab aktiivselt mööda kaheahelalist DNA molekuli ühest otsast teise, jättes selle taha hargnenud dubleeritud "saba". Selle valgu sellise töö füüsikalised põhimõtted pole veel selged.
Kuid DNA ja selle üksikud funktsionaalsed piirkonnad, mis kannavad teavet valkude struktuuri kohta, ei osale ise otseselt valgumolekulide loomise protsessis. Esimene samm selle DNA ahelatesse salvestatud teabe realiseerimiseks on nn transkriptsiooniprotsess või "ümberkirjutamine". Selle protsessi käigus sünteesitakse ühel DNA ahelal, nagu maatriksil, keemiliselt sarnane polümeer, ribonukleiinhape (RNA). RNA molekul on üheahelaline, mille monomeerideks on nelja tüüpi ribonukleotiidid, mida peetakse nelja tüüpi DNA desoksüribonukleotiidide kergeks modifikatsiooniks. Saadud RNA ahela nelja tüüpi ribonukleotiidide paigutuse järjestus kordab täpselt ühe kahest DNA ahelast vastavate desoksüribonukleotiidide järjestuse järjestust. Nii kopeeritakse geenide nukleotiidne järjestus RNA molekulide kujul, st. antud geeni struktuuris salvestatud informatsioon kopeeritakse täielikult RNA-sse. Igast geenist saab eemaldada suure, teoreetiliselt piiramatu arvu selliseid "koopiaid" – RNA molekule. Need molekulid, mis on paljudes koopiates ümber kirjutatud geenide "koopiateks" ja kannavad seetõttu geenidega sama teavet, hajuvad kogu rakus. Nad suhtlevad juba vahetult raku valke sünteesivate osakestega ja võtavad "isiklikult" osa valgumolekulide loomise protsessidest. Teisisõnu edastavad nad teavet selle salvestamise kohast selle realiseerimise kohta. Sellest lähtuvalt nimetatakse neid RNA-sid messengeriks (mRNA) või messengeriks (mRNA).
On leitud, et mRNA ahel sünteesitakse otse, kasutades matriitsina vastavat DNA piirkonda. Sünteesitud mRNA ahel kopeerib täpselt ühte kahest DNA ahelast oma nukleotiidjärjestuses (eeldusel, et uratsiil (U) RNA-s vastab selle derivaadile tümiinile (T) DNA-s). See toimub sama struktuurse komplementaarsuse põhimõtte alusel, mis määrab DNA reduplikatsiooni (joonis 18). Selgus, et kui mRNA sünteesitakse DNA-l rakus, kasutatakse mRNA ahela moodustamisel mallina ainult ühte DNA ahelat. Siis vastab iga selle DNA ahela G C-le ehitatavas RNA ahelas, igale DNA ahela C-le - G-le RNA ahelas, igale DNA ahela T-le - A-le RNA ahelas ja igale DNA-ahela A-le. - Y RNA ahelas. Selle tulemusena on saadud RNA ahel rangelt komplementaarne DNA matriitsi ahelaga ja seega nukleotiidjärjestuselt identne (eeldades, et T = Y) teise DNA ahelaga. Seega “kirjutatakse” info DNA-st RNA-ks, s.t. transkriptsioon. "Ümberkirjutatud" nukleotiidide kombinatsioonid RNA ahelas määravad juba otseselt nende poolt kodeeritud vastavate aminohapete paigutuse valguahelas.
Siin, nagu DNA reduplikatsiooni puhul, peab transkriptsiooniprotsessi üks olulisemaid aspekte olema selle ensümaatiline iseloom. DNA, mis on selles protsessis matriitsiks, määrab täielikult nukleotiidide paigutuse sünteesitud mRNA ahelas, kogu saadud RNA spetsiifilisuse, kuid protsessi ise viib läbi spetsiaalne valk - ensüüm. Seda ensüümi nimetatakse RNA polümeraasiks. Selle molekulil on keeruline organisatsioon, mis võimaldab tal aktiivselt liikuda mööda DNA molekuli, sünteesides samaaegselt ühe DNA ahelaga komplementaarset RNA ahelat. Maatriksina toimivat DNA molekuli ei tarbita ja see ei muutu, jäädes oma esialgsele kujule ja on alati valmis selliseks piiramatu arvu "koopiate" - mRNA - ümberkirjutamiseks. Nende mRNA-de voog DNA-st ribosoomidesse kujutab endast infovoogu, mis tagab raku valke sünteesiva aparaadi, selle ribosoomide terviku programmeerimise.
Seega kirjeldab skeemi vaadeldav osa DNA-st infovoogu mRNA molekulide kujul valke sünteesivate intratsellulaarsete osakesteni. Nüüd pöördume teistsuguse voolu poole – selle materjali voolu poole, millest valk tuleb luua. Valgumolekuli elementaarühikud - monomeerid - on aminohapped, mida on umbes 20. Valgu molekuli loomiseks (sünteesiks) peavad rakus olevad vabad aminohapped olema kaasatud vastavasse valku sünteesivasse voolu sisenevasse voolu. osakesi ja juba seal on nad paigutatud ahelasse teatud ainulaadsel viisil, mille dikteerib messenger RNA. See aminohapete kaasamine ehitusmaterjal valgu loomiseks - toimub vabade aminohapete kinnitamisega suhteliselt väikese suurusega spetsiaalsetele RNA molekulidele. Nendel RNA-del, mille ülesanne on kinnitada neile vabu aminohappeid, kuigi nad ei ole informatiivsed, on erinev - adapter - funktsioon, mille tähendust nähakse hiljem. Aminohapped on kinnitatud ülekande-RNA (tRNA) väikeste ahelate ühte otsa, üks aminohape RNA molekuli kohta. Iga sellise aminohappe jaoks rakus on spetsiifilised adapter-RNA molekulid, mis seovad ainult neid aminohappeid. Sellisel RNA-le riputatud kujul sisenevad aminohapped valke sünteesivatesse osakestesse.
Valkude biosünteesi protsessi keskseks momendiks on nende kahe rakusisese voolu – infovoo ja materjalivoo – sulandumine raku valke sünteesivates osakestes. Neid osakesi nimetatakse ribosoomideks. Ribosoomid on molekulaarsete mõõtmetega ultramikroskoopilised biokeemilised "masinad", kus vastavalt messenger-RNA-s sisalduvale plaanile pannakse sissetulevatest aminohappejääkidest kokku spetsiifilised valgud. Kuigi joonisel fig. 19 näitab ainult ühte osakest, iga rakk sisaldab tuhandeid ribisid. Ribosoomide arv määrab valgu sünteesi üldise intensiivsuse rakus. Ühe ribosoomiosakese läbimõõt on umbes 20 nm. Omal moel keemiline olemus ribosoom - ribonukleoproteiin: koosneb spetsiaalsest ribosomaalsest RNA-st (see on meile teadaolev RNA kolmas klass lisaks info- ja adapter-RNA-le) ja struktuursetest ribosomaalsetest valgu molekulidest. Üheskoos moodustab see mitmekümnest makromolekulist koosnev kombinatsioon ideaalselt organiseeritud ja töökindla “masina”, millel on võime lugeda mRNA ahelas sisalduvat informatsiooni ja realiseerida see kindla struktuuriga valmis valgumolekuli kujul. Kuna protsessi olemus seisneb selles, et 20 erineva aminohappe lineaarne paigutus valguahelas on unikaalselt määratud nelja erineva nukleotiidi paigutusega keemiliselt täiesti erineva polümeeri – nukleiinhappe (mRNA) ahelas, siis see protsess, mis esineb ribosoomis, seda nimetatakse tavaliselt "tõlkeks" või "tõlkeks" - translatsioon otsekui nukleiinhappeahelate neljatähelisest tähestikust valgu (polüpeptiid) ahelate kahekümnetäheliseks tähestikuks. Nagu näete, on kõik kolm tõlkeprotsessi kaasatud. kuulus klass RNA: messenger RNA, mis on translatsiooni objekt; ribosomaalne RNA, mis täidab valke sünteesiva ribonukleoproteiini osakese - ribosoomi - organisaatori rolli; ja adapter-RNA-d, mis täidavad translaatori funktsiooni.
|
Riis. 19. Funktsioneeriva ribosoomi skeem |

Valgu sünteesi protsess algab aminohappeühendite moodustumisega adapter-RNA molekulidega ehk tRNA-dega. Sel juhul toimub kõigepealt aminohappe energeetiline "aktiveerimine" selle ensümaatilise reaktsiooni tõttu adenosiintrifosfaadi (ATP) molekuliga ja seejärel ühendatakse "aktiveeritud" aminohape suhteliselt lühikese tRNA ahela otsaga, samas kui aktiveeritud aminohappe keemilise energia juurdekasv salvestatakse energia kujul keemiline side aminohapete ja tRNA vahel.
Samal ajal lahendatakse teine ülesanne. Fakt on see, et reaktsiooni aminohappe ja tRNA molekuli vahel viib läbi ensüüm, mida nimetatakse aminoatsüül-tRNA süntetaasiks. Igal 20 aminohappel on oma spetsiaalsed ensüümid, mis viivad reaktsiooni läbi ainult selle aminohappe osalusel. Seega on olemas vähemalt 20 ensüümi (aminoatsüül-tRNA süntetaas), millest igaüks on spetsiifiline ühe konkreetse aminohappe suhtes. Kõik need ensüümid võivad reageerida mitte ühegi tRNA molekuliga, vaid ainult nendega, mille ahelas on rangelt määratletud nukleotiidide kombinatsioon. Seega, kuna on olemas hulk selliseid spetsiifilisi ensüüme, mis eristavad ühelt poolt aminohappe olemust ja teiselt poolt tRNA nukleotiidjärjestust, on kõik 20 aminohapet "määratud". ainult teatud tRNA-dele, millel on antud iseloomulik nukleotiidide kombinatsioon.
Skemaatiliselt on mõned valkude biosünteesi protsessi hetked, nii palju kui me neid täna esitame, toodud joonisel fig. 19. Siin on esiteks näha, et ribosoomiga on ühendatud RNA-molekul ehk, nagu öeldakse, on ribosoomi “programmeeritud” messenger-RNA poolt. Igal ajahetkel paikneb ainult suhteliselt lühike mRNA ahela segment otse ribosoomis endas. Kuid just see segment võib ribosoomi osalusel suhelda adapter-RNA molekulidega. Siingi mängib suurt rolli komplementaarsuse põhimõte.
See on mehhanismi selgitus, miks mRNA ahela antud kolmik vastab rangelt määratletud aminohappele. Adapter-RNA (tRNA) on vajalik vahelüli ehk adapter, kui iga aminohape "tunneb ära" oma tripleti mRNA-l.
Joonisel fig. Joonisel 19 on näha, et lisaks tRNA molekulile koos kinnitatud aminohappega on ribosoomis veel üks tRNA molekul. Kuid erinevalt ülalkirjeldatud tRNA molekulist on see tRNA molekul kinnitatud oma otsaga sünteesiprotsessis oleva valgu (polüpeptiid) ahela otsa. See asend peegeldab valgumolekuli sünteesi ajal ribosoomides toimuvate sündmuste dünaamikat. Seda dünaamikat võib ette kujutada järgmiselt. Alustame mõne vahepunktiga, mis on näidatud joonisel fig. 19 ja seda iseloomustab juba ehitama hakatud valguahela olemasolu, sellega seotud tRNA, mis on just sisenenud ribosoomi ja on seotud uue tRNA molekuli kolmikuga koos vastava aminohappega. Ilmselt viib juba tRNA molekuli kinnitumine ribosoomi antud kohas asuva mRNA kolmiku külge sellise vastastikuse orientatsiooni ja tiheda kontakti aminohappejäägi ja ehitatava valguahela vahel, et kovalentne side. Ühendus toimub nii, et konstrueeritava valguahela ots (joonisel 19 kinnitatud tRNA-ga) kantakse sellelt tRNA-lt üle saabuva aminoatsüül-tRNA aminohappejäägile. Selle tulemusena on "õige" tRNA, mis on täitnud "doonori" rolli, vabaks ja valgu ahel- kantakse üle "vastuvõtjale", s.o. "vasakul" (sissetulev) aminoatsüül-tRNA. Selle tulemusena pikeneb valguahel ühe aminohappe võrra ja kinnitub "vasakpoolse" tRNA külge. Sellele järgneb “vasakpoolse” tRNA ülekandmine koos sellega seotud mRNA nukleotiidide kolmikuga paremale, seejärel nihkub siit välja endine “doonor” tRNA molekul ja lahkub ribosoomidest. Selle asemele ilmub uus tRNA koos ehitatava valguahelaga, mida pikendatakse ühe aminohappejäägi võrra ja mRNA ahel edeneb ribosoomi suhtes kolmiku võrra paremale. MRNA ahela ühe tripleti võrra paremale nihutamise tulemusena tekib ribosoomi järgmine vakantne kolmik (UUU), millele lisandub komplementaarsel põhimõttel kohe vastav tRNA koos aminohappega (fenüülalanüül-tRNA). See põhjustab taas kovalentse (peptiid) sideme moodustumist ehitatava valguahela ja fenüülalaniini jäägi vahel ning pärast seda liigub mRNA ahel ühe tripleti võrra paremale koos kõigi sellest tulenevate tagajärgedega jne. Nii tõmmatakse järjestikku, kolmik-tripleti haaval info-RNA ahel läbi ribosoomi, mille tulemusena “loetakse” mRNA ahelat ribosoomi poolt tervikuna algusest lõpuni. Samaaegselt ja koos sellega toimub järjestikuste aminohapete kaupa valguahela kogunemine. Vastavalt sellele sisenevad aminohapetega tRNA molekulid üksteise järel ribosoomi ja ilma aminohapeteta tRNA molekulid väljuvad. Leides end väljaspool ribosoomi lahuses, ühinevad vabad tRNA molekulid taas aminohapetega ja kannavad need uuesti ribosoomi, tsirkuleerides seega tsükliliselt ilma hävimise ja muutumiseta.
Bioloogilistes järjestustes sisalduv teave
Biopolümeerid on (bioloogilised) polümeerid, mida sünteesivad elusolendid. DNA, RNA ja valgud on lineaarsed polümeerid, st iga monomeer, mida need sisaldavad, ühineb vähemalt kahe teise monomeeriga. Monomeeride jada kodeerib informatsiooni, mille ülekandereegleid kirjeldab keskne dogma. Teave edastatakse suure täpsusega, deterministlikult ja ühte biopolümeeri kasutatakse mallina teise polümeeri kokkupanemisel järjestusega, mis on täielikult määratud esimese polümeeri järjestusega.
Universaalsed viisid bioloogilise teabe edastamiseks
Elusorganismides on kolme tüüpi heterogeenseid, st koosnevad erinevatest polümeeri monomeeridest - DNA, RNA ja valk. Teabe edastamist nende vahel saab läbi viia 3 × 3 = 9 viisil. Keskne dogma jagab need 9 teabeedastuse tüüpi kolme rühma:
- Üldine – leidub enamikus elusorganismides;
- Spetsiaalne - leitakse erandkorras viirustes ja genoomi mobiilsetes elementides või bioloogilise katse tingimustes;
- Tundmatu – ei leitud.
DNA replikatsioon (DNA → DNA)
DNA on peamine teabe edastamise viis elusorganismide põlvkondade vahel, seega on DNA täpne dubleerimine (replikatsioon) väga oluline. Replikatsiooni viib läbi valkude kompleks, mis kerivad lahti kromatiini ja seejärel topeltheeliksi. Pärast seda loovad DNA polümeraas ja sellega seotud valgud mõlemale ahelale identse koopia.
Transkriptsioon (DNA → RNA)
Transkriptsioon on bioloogiline protsess, mille tulemusena kopeeritakse DNA segmendis sisalduv informatsioon sünteesitud mRNA molekulile. Transkriptsioon viiakse läbi transkriptsioonifaktorite ja RNA polümeraasi abil. Eukarüootses rakus redigeeritakse sageli esmast transkripti (pre-mRNA). Seda protsessi nimetatakse splaissimiseks.
Skemaatiline diagramm geneetilise teabe realiseerumisest pro- ja eukarüootides.PROKARÜOODID. Prokarüootides ei ole ribosoomi valgu süntees (translatsioon) transkriptsioonist ruumiliselt eraldatud ja see võib toimuda isegi enne mRNA sünteesi lõpetamist RNA polümeraasi poolt. Prokarüootsed mRNA-d on sageli polütsistroonilised, mis tähendab, et need sisaldavad mitut sõltumatut geeni.
EUKARÜOODID. eukarüootne mRNA sünteesitakse prekursorina, pre-mRNA-na, mis seejärel läbib keerulise etapiviisilise küpsemise – töötlemise, sealhulgas korgistruktuuri kinnitamise molekuli 5 "otsa, mitmekümne adeniinijäägi kinnitumist selle 3" otsa (polüadenüülimine), ebaoluliste sektsioonide - intronite - lõhustamine ja oluliste sektsioonide - eksonite - üksteisega ühendamine (splaissimine). Sel juhul võib toimuda sama pre-mRNA eksonite ühendamine erinevatel viisidel, mis viib erinevate küpsete mRNA-de ja lõpuks erinevate valguvariantide moodustumiseni (alternatiivne splaissimine). Ainult edukalt töödeldud mRNA eksporditakse tuumast tsütoplasmasse ja osaleb translatsioonis.
Tõlge (RNA → valk)
RNA replikatsioon (RNA → RNA)
RNA replikatsioon – RNA ahela kopeerimine selle komplementaarseks RNA ahelaks, kasutades ensüümi RNA-sõltuvat RNA polümeraasi. Üheahelalist (näiteks pikornaviirused, mis hõlmavad suu- ja sõrataudiviirust) või kaheahelalist RNA-d sisaldavad viirused paljunevad sarnasel viisil.
Valgu otsene translatsioon DNA matriitsile (DNA → valk)
Elus translatsiooni on demonstreeritud E. coli rakuekstraktides, mis sisaldasid ribosoome, kuid mitte mRNA-d. Sellised ekstraktid sünteesisid süsteemi sisestatud DNA-st valke ja antibiootikum neomütsiin tugevdas seda toimet.
Epigeneetilised muutused
Epigeneetilised muutused on muutused geenide ekspressioonis, mida ei põhjusta geneetilise informatsiooni muutused (mutatsioonid). Epigeneetilised muutused tekivad geeniekspressiooni taseme muutmise, st nende transkriptsiooni ja/või translatsiooni tulemusena. Enim uuritud epigeneetilise regulatsiooni tüüp on DNA metüülimine DNA metüültransferaasi valkude abil, mis viib metüleeritud geeni ajutise, elust sõltuva inaktiveerimiseni. Kuna aga DNA molekuli primaarstruktuur ei muutu, ei saa seda erandit pidada tõeliseks näiteks informatsiooni ülekandest valgust DNA-sse.
prioonid
Prioonid on valgud, mis eksisteerivad kahel kujul. Valgu üks vorme (konformatsioon) on funktsionaalne, tavaliselt vees lahustuv. Teine vorm moodustab vees lahustumatuid agregaate, sageli molekulaarsete polümeertorude kujul. Selles konformatsioonis olev monomeer – valgumolekul – on võimeline ühinema teiste sarnaste valgumolekulidega, muutes need teiseks, prioonilaadseks konformatsiooniks. Seentes võivad sellised molekulid olla päritud. Kuid nagu DNA metüülimise puhul, jääb valgu esmane struktuur sel juhul samaks ja nukleiinhapetele infot ei edastata.
Mõiste "dogma" ajalugu
originaaltekst(Inglise)
Minu arvamus oli, et dogma oli idee, mille kohta puudusid mõistlikud tõendid. Näete?!" Ja Crick möirgas rõõmust. "Ma lihtsalt ei teadnud, mida dogma tähendab. Ja sama hästi oleksin võinud seda nimetada "keskseks hüpoteesiks" või - teate. Mida ma tahtsin ütleme, et dogma oli lihtsalt tabav lause
Lisaks kirjutas Crick oma autobiograafilises raamatus "What a Mad Pursuit" sõna "dogma" valikust ja selle valiku põhjustatud probleemidest:
"Ma kahtlustan, et nimetasin seda ideed keskseks dogmaks kahel põhjusel. Jadahüpoteesis olen juba kasutanud sõna hüpotees, pealegi tahtsin vihjata, et see uus oletus on kesksem ja tugevam ... Nagu selgub, tekitas dogma mõiste kasutamine rohkem tüli, kui see väärt oli ... Paljud aastaid hiljem ütles Jacques Monod mulle, et ilmselt ma ei saanud aru, mida mõeldakse sõna dogma all, mis tähendab osa usust, mis ei allu kahtlustele. Ma olin selle sõna tähenduse pärast ähmaselt kartlik, kuid kuna ma uskusin, et kõigil usulistel veendumustel pole alust, siis kasutasin seda sõna nii, nagu ma sellest aru sain, ja mitte enamik teisi inimesi, rakendades seda suurejoonelisele hüpoteesile, et hoolimata usaldusest see inspireeritud, põhines väikesel hulgal otsestel katseandmetel.
originaaltekst(Inglise)
Ma kahtlustan, et nimetasin seda ideed keskseks dogmaks kahel põhjusel. Olin juba kasutanud ilmset sõnahüpoteesi jadahüpoteesis ja lisaks tahtsin väita, et see uus oletus on kesksem ja võimsam. ... Nagu selgus, tekitas sõna dogma kasutamine peaaegu rohkem probleeme, kui see väärt oli... Palju aastaid hiljem juhtis Jacques Monod mulle tähelepanu, et ma ei paistnud aru saavat ja sõna dogma õige kasutamine, mis on veendumus, milles ei saa kahelda. Ma mõistsin seda ebamäärasel viisil, kuid kuna arvasin, et kõik religioossed tõekspidamised on alusetud, kasutasin seda sõna nii, nagu ma ise sellest arvasin, mitte nagu enamik maailmast, ja rakendasin seda lihtsalt suure hüpoteesi jaoks. millel, olgugi usutav, oli vähe otsest eksperimentaalset tuge.
Vaata ka
Märkmed
Lingid
- B. J. McCarthy, J. J. Holland. Denatureeritud DNA otsese mallina in vitro Valkude süntees // PNAS. - 1965. - T. 54. - S. 880-886.
- Werner, E. Genoomi semantika, In Silico mitmerakulised süsteemid ja keskne dogma // FEBSi kirjad. - 2005. - V. 579. - S. 1779-1782. PMID 15763551
- Horace Freeland Judson. 6. peatükk: Minu arvamus oli, et dogma oli idee, mille kohta puudusid mõistlikud tõendid. Sa näed?! // Loomise kaheksas päev: Bioloogia revolutsiooni tegijad (25. aastapäeva väljaanne). - 1996.
Molekulaarbioloogia keskne dogma
Raku tuuma struktuur
Rakkude fraktsioneerimine.Tänapäeval võimaldab fraktsioneerimine saada peaaegu kõiki raku organelle ja struktuure: tuumad, nukleoolid, kromatiini, tuumamembraanid, plasmamembraan, endoplasmaatilise retikulumi vakuoolid jne.
Spetsiaalsed meetodid
Enne rakufraktsioonide saamist rakud hävitatakse homogeniseerimise teel. Lisaks eraldatakse homogenaatidest fraktsioonid. Eraldustsentrifuugimine on peamine meetod rakustruktuuride eraldamiseks. See põhineb asjaolul, et raskemad osakesed settivad kiiremini tsentrifuugitoru põhja.
Madalatel kiirendustel (1-3 tuhat g) settivad tuumad ja terved rakud varem, 15-30 tuhande g juures, suuremad osakesed või makrosoomid, mis koosnevad mitokondritest, väikestest plastiididest, peroksisoomidest, lüsosoomidest jne, settivad 50 tuhande g juures. , mikrosoomid, raku vakuolaarsüsteemi fragmendid, settivad. Segatud alamfraktsioonide uuesti tsentrifuugimisel eraldatakse puhtad fraktsioonid. Fraktsioonide peenemaks eraldamiseks kasutatakse sahharoosi tiheduse gradiendi tsentrifuugimist. Üksikute rakukomponentide saamine võimaldab uurida nende biokeemiat ja funktsionaalseid omadusi, luua rakuvabad süsteemid, näiteks ribosoomide jaoks, mis suudavad sünteesida valke vastavalt eksperimenteerija määratud RNA-le, või raku supramolekulaarsete struktuuride taasloomiseks.
Majutatud aadressil ref.rf
Sellised tehissüsteemid aitavad uurida rakus toimuvaid peeneid protsesse.
meetod rakutehnoloogia. Pärast eritöötlust võivad erinevad elusrakud omavahel sulanduda ja moodustada kahetuumalise raku ehk heterokarüoni. Heterokarüonid, eriti need, mis on moodustunud lähedastest rakkudest (näiteks hiired ja hamstrid), võivad siseneda mitoosi ja tekitada tõelisi hübriidrakke. Teised tehnikad võimaldavad konstrueerida rakke erineva päritoluga tuumadest ja tsütoplasmast.
Tänapäeval kasutatakse rakutehnoloogiat laialdaselt mitte ainult eksperimentaalbioloogias, vaid ka biotehnoloogias. Näiteks monoklonaalsete antikehade saamisel.
Rakul on tohutult palju erinevaid funktsioone, peamisteks töömehhanismideks nende funktsioonide täitmiseks on valgud või nende kompleksid teiste bioloogiliste makromolekulidega. Peaaegu kõik sünteesi-, lagunemis-, erinevate valkude, nukleiinhapete, lipiidide, süsivesikute ümberkorraldamise protsessid toimuvad ensüümvalkude osalusel. Kontraktsiooni, mis viib rakkude liikuvuseni või ainete ja struktuuride liikumiseni rakkude sees, teostavad ka spetsiaalsed kontraktiilsed valgud. Paljud raku reaktsioonid vastuseks välisteguritele (viirused, hormoonid, võõrvalgud jne) saavad alguse nende tegurite koostoimest spetsiaalsete raku retseptorvalkudega.
Valgud on peaaegu kõigi rakustruktuuride põhikomponendid.
Majutatud aadressil ref.rf
Iga üksiku valgu struktuur on rangelt spetsiifiline, mis väljendub nende esmase struktuuri spetsiifilisuses - aminohapete järjestuses piki polüpeptiidi, valguahelat. Sellise korrektsuse valguahelas üheselt mõistetava aminohappejärjestuse reprodutseerimisel määrab selle geenipiirkonna DNA struktuur, mis lõppkokkuvõttes vastutab selle valgu struktuuri ja sünteesi eest. See seisukoht on molekulaarbioloogia või selle ʼʼdogmaʼʼ põhipostulaat. Lisaks rõhutab keskne dogma teabe ühesuunalist edastamist: ainult DNA-st valku (DNA ® mRNA ® valk) ja eitab tagasiteed - valgust nukleiinhappeni.
Praeguste teadmiste põhjal on valkude biosüntees järgmine skemaatiline diagramm.
peamist rolli valkude spetsiifilise struktuuri määramisel kuulub DNA. Kahest põimunud polümeeriahelast koosnev DNA molekul on lineaarne struktuur, mille monomeerideks on nelja tüüpi desoksüribonukleotiidid, mille vaheldumine või järjestus piki ahelat on ainulaadne ja spetsiifiline iga DNA molekuli ja iga selle lõigu jaoks. Iga valgu sünteesi eest vastutab DNA molekuli konkreetne piirkond. DNA molekuli osa, mis sisaldab kogu teavet ühe vastava valgu struktuuri kohta. nimetatakse tsistroniks. Tänapäeval peetakse tsistroni mõistet samaväärseks geeni mõistega.
On teada, et erinevalt teistest valke sünteesiva aparaadi komponentidest on eukarüootsete organismide DNA asukohaks rakkudes raku tuum. Madalamates (prokarüootsetes) organismides, millel ei ole hästi moodustunud rakutuum, eraldatakse DNA ka ülejäänud protoplasmast ühe või mitme kompaktse nukleotiidi kujul.
DNA makromolekulaarse struktuuri juur on niinimetatud komplementaarsuse põhimõte. See tähendab, et kahe põimunud DNA ahela vastassuunalised nukleotiidid täiendavad üksteist oma ruumilise struktuuriga. Sellised komplementaarsed - komplementaarsed - nukleotiidipaarid on A-T paar (adeniin-tüümiin) ja G-C paar (guaniin-tsütosiin).
Uute DNA molekulide süntees rakus toimub ainult olemasolevate DNA molekulide baasil. Sel juhul hakkavad algse DNA molekuli kaks ahelat ühest otsast lahknema ja igal eraldatud üheahelalisel lõigul hakkab teine ahel söötmes sisalduvatest vabadest nukleotiididest kogunema rangelt kooskõlas täiendavuse põhimõtet. Igas "tütar" DNA molekulis pärineb üks ahel täielikult originaalist ja teine on äsja sünteesitud.
Tuleb rõhutada, et täpse paljunemise potentsiaal on omane DNA enda kaheahelalisele komplementaarsele struktuurile ja selle avastamine on bioloogia üks peamisi saavutusi.
Sünteesi - DNA reprodutseerimise protsessi läbiviimiseks vastavalt ülalkirjeldatud skeemile on vajalik spetsiaalse ensüümi, mida nimetatakse DNA polümeraasiks, aktiivsus. Just see ensüüm liigub järjestikku DNA molekuli ühest otsast teise, kahe ahela eraldamise protsess koos nende vabade nukleotiidide samaaegse polümerisatsiooniga vastavalt komplementaarsele põhimõttele.
Järelikult määrab DNA nagu maatriks ainult nukleotiidide järjestuse sünteesitud ahelates ja protsessi ise viib läbi valk. DNA ja selle üksikud funktsionaalsed piirkonnad, mis kannavad teavet valkude struktuuri kohta, ei osale ise otseselt valgumolekulide loomise protsessis. Esimene samm selle teabe realiseerimise suunas on nn transkriptsiooniprotsess ehk ʼʼümberkirjutamineʼʼ. Selles protsessis sünteesitakse DNA ahelas nagu maatriksil keemiliselt seotud polümeer, ribonukleiinhape (RNA). RNA molekul on üheahelaline, mille monomeerideks on nelja tüüpi ribonukleotiidid. Saadud RNA ahela nelja tüüpi ribonukleotiidide paigutuse järjestus kordab täpselt ühe kahest DNA ahelast vastavate desoksüribonukleotiidide järjestuse järjestust. Tänu sellele kopeeritakse selle geeni struktuuris salvestatud teave täielikult RNA-sse. Igast geenist saab eemaldada teoreetiliselt piiramatu arvu ʼʼkoopiaidʼʼ – RNA molekule. RNA molekulid astuvad suhtlemisse raku valke sünteesivate osakestega ja osalevad otseselt valgumolekulide sünteesis. Teisisõnu edastavad nad teavet selle salvestamise kohtadest selle rakendamise kohtadesse. Seetõttu nimetatakse neid RNA-sid messenger- või messenger-RNA-deks, lühendatult mRNA-ks või mRNA-ks.
Sünteesitud messenger-RNA ahel kasutab matriitsina otse vastavat DNA piirkonda. Sel juhul kopeerib sünteesitud mRNA ahel täpselt ühte kahest DNA ahelast oma nukleotiidjärjestuses (uratsiil (U) RNA-s vastab selle derivaadile tümiinile (T) DNA-s). Kõik toimub sama komplementaarsuse põhimõtte alusel, mis määrab DNA reduplikatsiooni. Selle tulemusena toimub teabe ʼʼümberkirjutamineʼʼʼ või transkriptsioon DNA-st RNA-sse. RNA nukleotiidide ʼʼÜmberkirjutatudʼʼ kombinatsioonid määravad juba otseselt nende poolt kodeeritud aminohapete paigutuse valguahelas.
Kuidas nüüd valku tehakse? Teatavasti on valgumolekuli monomeeride tüübid aminohapped, mida on 20 erinevat sorti. Igat tüüpi aminohapete jaoks rakus on spetsiifilised adapter-RNA molekulid, mis kinnitavad ainult seda tüüpi aminohappeid. RNA-l külastatud kujul sisenevad aminohapped valke sünteesivatesse osakestesse - ribosoomidesse ja juba seal, sõnumitooja RNA dikteerimisel, paigutatakse nad sünteesitava valgu ahelasse.
Valkude biosünteesis on põhiline kahe rakusisese voo liitmine ribosoomides – infovoog ja materjalivoog. Ribosoomid on molekulaarsete mõõtmetega biokeemilised "masinad", milles saabuvatest aminohappejääkidest pannakse kokku spetsiifilised valgud vastavalt messenger-RNA-s sisalduvale plaanile. Iga rakk sisaldab tuhandeid ribsoome, valgusünteesi intensiivsuse määrab nende arv rakus. Oma keemilise olemuse järgi kuulub ribosoom ribonukleoproteiinide hulka ja koosneb spetsiaalsest ribosomaalsest RNA-st ja ribosomaalsetest valgu molekulidest. Ribosoomidel on võime lugeda mRNA ahelas sisalduvat teavet ja realiseerida seda valmis valgu molekuli kujul. Protsessi olemus seisneb selles, et 20 tüüpi aminohapete lineaarne paigutus valguahelas määratakse nelja tüüpi nukleotiidide paigutusega täiesti erineva polümeeri - nukleiinhappe (mRNA) ahelas. Sel põhjusel nimetatakse seda ribosoomis toimuvat protsessi tavaliselt "translatsiooniks" või "translatsiooniks" - translatsiooniks nukleiinhappeahelate 4-tähelisest tähestikust valgu (polüpeptiid) ahelate 20-täheliseks tähestikuks. Selles translatsiooniprotsessis osalevad kõik kolm teadaolevat RNA klassi: messenger RNA, mis on translatsiooni objektiks, ribosoomi RNA, mis täidab ribosoomi organisaatori rolli, ja adapter-RNA, mis toimib translaatorina.
Valgu sünteesi protsess algab aminohappeühendite moodustumisega adapter-RNA molekulidega. Sel juhul toimub kõigepealt aminohappe energia ʼʼaktiveerimineʼʼ tänu selle ensümaatilisele reaktsioonile adenosiintrifosfaadi (ATP) molekuliga ja seejärel ühendatakse "aktiveeritud" aminohape suhteliselt lühikese tRNA ahela otsaga, samal ajal aktiveeritud aminohappe keemiline energia salvestub aminohappe ja tRNA vahelise keemilise sideme energia kujul.
Tuleb lisada, et reaktsiooni aminohappe ja tRNA molekuli vahel viib läbi ensüüm aminoatsüül-tRNA süntetaas. Igal 20 aminohappel on oma ensüümid, mis viivad reaktsiooni läbi ainult selle aminohappe osalusel.
Molekulaarbioloogia keskne dogma on mõiste ja tüübid. Kategooria "Molekulaarbioloogia keskdogma" klassifikatsioon ja tunnused 2017, 2018.
Maatriksi biosünteesi põhifiguuriks on nukleiinhapped RNA ja DNA. Need on polümeersed molekulid, mis sisaldavad viit tüüpi lämmastikaluseid, kahte tüüpi pentoose ja fosforhappejääke. Nukleiinhapetes võivad lämmastiku alused olla puriin (adeniin, guaniin) ja pürimidiin (tsütosiin, uratsiil (ainult RNA-s), tümiin (ainult DNA-s)). Sõltuvalt süsivesikute struktuurist, ribonukleiinhapped - sisaldavad riboosi (RNA) ja desoksüribonukleiinhapped- sisaldavad desoksüriboosi (DNA).
Mõiste " maatriksi biosünteesid "tähendab raku võimet sünteesida polümeerseid molekule, nagu nukleiinhapped Ja oravad, põhineb malli-maatriksil. See tagab kõige keerukama struktuuri täpse ülekande olemasolevatelt molekulidelt äsja sünteesitud molekulidele.
Molekulaarbioloogia põhipostulaat
Enamikul juhtudel edastamine pärilikku teavet emarakust tütarrakku toimub DNA abil (replikatsioon). Geneetilise informatsiooni kasutamiseks raku enda poolt on vaja RNA-d, mis moodustub DNA matriitsil (transkriptsioon). Lisaks osaleb RNA otseselt raku struktuuri ja aktiivsust tagavate valgumolekulide sünteesi (translatsiooni) kõigis etappides.
Eeltoodu põhjal Molekulaarbioloogia keskne dogma, mille kohaselt toimub geneetilise informatsiooni ülekanne ainult nukleiinhapetest (DNA ja RNA). Teabe vastuvõtjaks võib olla teine nukleiinhape (DNA või RNA) ja valk.
Molekulaarsete keskne dogma bioloogia - reegel, mis üldistab looduses vaadeldava geneetilise teabe rakendamist: teave edastatakse nukleiinhapetelt valgule, kuid mitte vastupidises suunas. Reegli sõnastas 1958. aastal Francis Crick ja viidi 1970. aastal vastavusse selleks ajaks kogutud andmetega. Geneetilise informatsiooni ülekanne DNA-st RNA-sse ja RNA-st valku on universaalne erandita kõikidele rakulistele organismidele ning on makromolekulide biosünteesi aluseks. Genoomi replikatsioon vastab DNA → DNA informatsioonilisele üleminekule. Looduses toimuvad ka üleminekud RNA → RNA ja RNA → DNA (näiteks mõnel viirusel), samuti molekulist molekuli edasi kanduvate valkude konformatsiooni muutus. Transkriptsioon ja tõlkimine. Tavaliselt saab kogu transkriptsiooni ja translatsiooni protsessi kuvada järgmisel diagrammil: Transkriptsioon on DNA-sse salvestatud teabe taasesitamise protsess üheahelalise molekuli ja RNA kujul (messenger RNA, mis edastab teavet valgu struktuuri kohta rakutuum raku tsütoplasmasse ribosoomideks). See protsess avaldub DNA matriitsist pärineva molekuli ja RNA sünteesis. Molekul ja RNA koosnevad nukleotiididest, millest igaüks sisaldab fosforhappejääki, suhkrut, riboosi ja ühte neljast lämmastikku sisaldavad alused(T-tuliini asemel A, G, C ja U-uratsiil). Süntees ja RNA põhineb komplementaarsuse põhimõttel, s.o. DNA ühes ahelas on A vastu Y ja RNA ning G vastu DNA-s - C ja RNA (vt joonis Transkriptsioon - eelmisel leheküljel), seega on RNA DNA või selle teatud lõigu komplementaarne koopia. ja sisaldab teavet, mis kodeerib aminohapet või valku. Iga aminohapet DNA-s ja RNA-s kodeerib 3 nukleotiidist koosnev järjestus, st. - triplett, mida nimetatakse koodoniks.Kui transkriptsioonis avaldub kahe molekuli äratundmine üksteise poolt vaid komplementaarsuse põhimõttes, siis translatsioonis lisaks komplementaarsusele (koodoni ja RNA ajutine kombinatsioon ning antikoodon RNA-st (transport RNA, mis toob valgusünteesiks vajalikud aminohapped sünteesikohta – ribosoomi – vt joon. Transkriptsioon) molekulaarne äratundmine avaldub ensüümi kodaasi abil aminohappe tRNA külge kinnitamise protsessis. et tRNA molekul koosneb peast, mis sisaldab anti-AOK tripletti, mis koosneb kolmest nukleotiidist koosnevast järjestusest, ja sabast, millel on kindel, mitut tüüpi tRNA antikosoneid eksisteerib, nii palju sabade vorme ja igal antikosoonil on oma saba kuju tRNA-s.Kui palju saba vorme eksisteerib, nii palju ensüümi kodaasi vorme, mis kinnitavad sabale aminohappeid, ja iga kodaasi kuju sobib ainult kujuga Seega kannab tRNA teavet endaga mitte. ainult antikosoonis olevate nukleotiidide n järjestuses, aga ka molekuli saba kujul. Ja peamine teabeedastus on siin valgu aminohapete järjestuse reprodutseerimine, mis kutsub esile valku ja RNA-d kodeeriva ensüümi.

| Eelmised materjalid: |