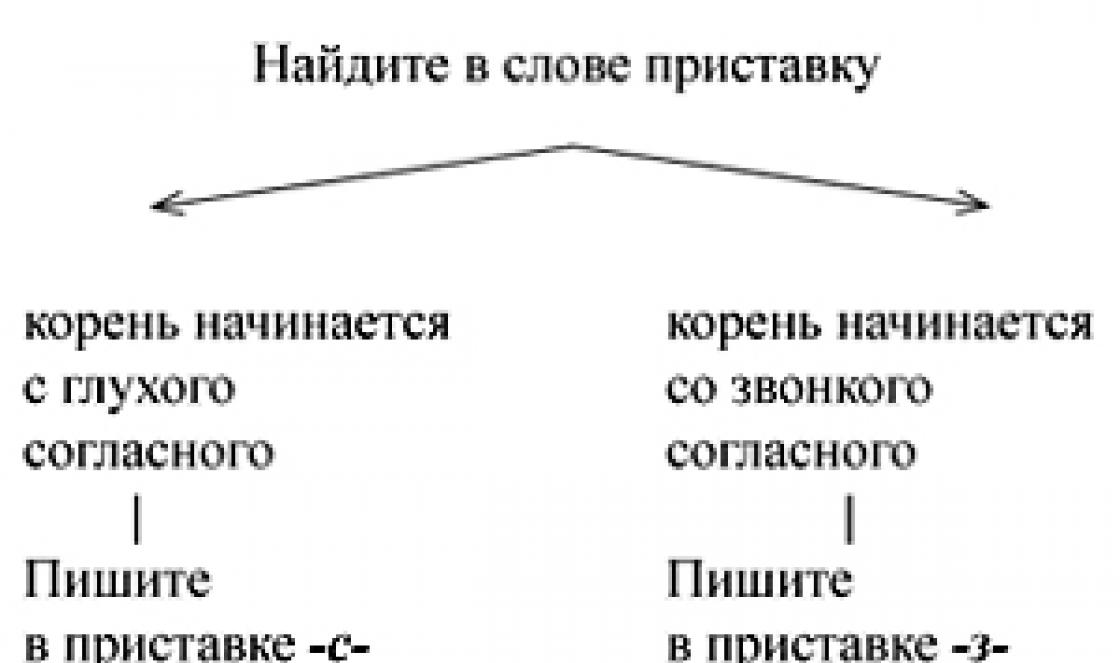
Որպեսզի հասկանանք ոչ միայն բջջի կառուցվածքային առանձնահատկությունների նշանակությունը, այլև, ամենակարևորը, հասկանալու նրա առանձին բաղադրիչների և ամբողջ բջջի ֆունկցիոնալ գործառույթները, որպեսզի համատեղենք բջջի մորֆոլոգիայի ուսումնասիրությունը նրա կառուցվածքի և աշխատանքի հիմնական կենսաքիմիական և գենետիկական առանձնահատկությունները, բջիջը ժամանակակից բջջային կենսաբանության դիրքերով ճշգրիտ ուսումնասիրելու համար անհրաժեշտ է գոնե համառոտ հիշել հիմնական մոլեկուլային կենսաբանական օրինաչափությունները, ևս մեկ անգամ հակիրճ անդրադառնալ կենտրոնական դոգմայի բովանդակությանը: մոլեկուլային կենսաբանություն.
Բջիջը որպես այդպիսին կատարում է բազմաթիվ տարբեր գործառույթներ: Ինչպես արդեն ասացինք, դրանցից մի քանիսը ընդհանուր բջջային են, որոշները՝ հատուկ, հատուկ բջջային տեսակներին բնորոշ։ Այս գործառույթների հիմնական աշխատանքային մեխանիզմները սպիտակուցներն են կամ դրանց բարդույթները այլ կենսաբանական մակրոմոլեկուլների հետ, ինչպիսիք են նուկլեինաթթուները, լիպիդները և պոլիսախարիդները: Օրինակ, հայտնի է, որ տարբեր նյութերի բջջում փոխադրման պրոցեսները՝ իոններից մինչև մակրոմոլեկուլներ, որոշվում են հատուկ սպիտակուցների կամ լիպոպրոտեինային համալիրների աշխատանքով, որոնք պլազմայի մաս են կազմում և այլն։ բջջային մեմբրաններ. Տարբեր սպիտակուցների սինթեզի, քայքայման, վերադասավորման գրեթե բոլոր գործընթացները, նուկլեինաթթուներ, լիպիդները, ածխաջրերը առաջանում են յուրաքանչյուր առանձին ռեակցիայի համար հատուկ սպիտակուց-ֆերմենտների գործունեության արդյունքում։ Առանձին կենսաբանական մոնոմերների, նուկլեոտիդների, ամինաթթուների սինթեզներ, ճարպաթթուներ, շաքարները և այլ միացությունները նույնպես իրականացվում են հսկայական քանակությամբ հատուկ ֆերմենտների՝ սպիտակուցների միջոցով։ Կծկումը, որը հանգեցնում է բջիջների շարժունակությանը կամ բջիջների ներսում նյութերի և կառուցվածքների շարժմանը, իրականացվում է նաև հատուկ կծկվող սպիտակուցների միջոցով։ Բջջային շատ ռեակցիաներ՝ ի պատասխան արտաքին գործոնների (վիրուսներ, հորմոններ, օտար սպիտակուցներ և այլն) սկսվում են այդ գործոնների փոխազդեցությամբ բջջային հատուկ ընկալիչների սպիտակուցների հետ։
Սպիտակուցները գրեթե բոլոր բջջային կառույցների հիմնական բաղադրիչներն են: Մի փունջ քիմիական ռեակցիաներբջիջի ներսում որոշվում է բազմաթիվ ֆերմենտներով, որոնցից յուրաքանչյուրը հանգեցնում է մեկ կամ մի քանի առանձին ռեակցիաների: Յուրաքանչյուր առանձին սպիտակուցի կառուցվածքը խիստ սպեցիֆիկ է, որն արտահայտվում է դրանց առաջնային կառուցվածքի յուրահատկությամբ՝ պոլիպեպտիդային, սպիտակուցային շղթայի երկայնքով ամինաթթուների հաջորդականությամբ։ Ավելին, այս ամինաթթուների հաջորդականության առանձնահատկությունն անվրեպ կրկնվում է այս բջջային սպիտակուցի բոլոր մոլեկուլներում:
Սպիտակուցային շղթայում ամինաթթուների միանշանակ հաջորդականությունը վերարտադրելու նման կոռեկտությունը որոշվում է այդ գենային շրջանի ԴՆԹ-ի կառուցվածքով, որն ի վերջո պատասխանատու է այս սպիտակուցի կառուցվածքի և սինթեզի համար: Այս գաղափարները ծառայում են որպես մոլեկուլային կենսաբանության հիմնական պոստուլատ՝ նրա «դոգմա»։ Ապագա սպիտակուցի մոլեկուլի մասին տեղեկատվությունը փոխանցվում է դրա սինթեզի վայրերին (ռիբոսոմների) միջնորդի միջոցով՝ սուրհանդակային ՌՆԹ (mRNA), որի նուկլեոտիդային բաղադրությունը արտացոլում է ԴՆԹ-ի գենային շրջանի կազմը և նուկլեոտիդային հաջորդականությունը: Ռիբոսոմում կառուցված է պոլիպեպտիդային շղթա, որի ամինաթթուների հաջորդականությունը որոշվում է mRNA-ում նուկլեոտիդների հաջորդականությամբ, դրանց եռյակների հաջորդականությամբ։ Այսպիսով, մոլեկուլային կենսաբանության կենտրոնական դոգման ընդգծում է տեղեկատվության միակողմանի փոխանցումը. միայն ԴՆԹ-ից սպիտակուց միջանկյալի՝ mRNA (ԴՆԹ → mRNA → սպիտակուց) օգնությամբ: Որոշ ՌՆԹ պարունակող վիրուսների համար տեղեկատվության փոխանցման շղթան կարող է հետևել ՌՆԹ → mRNA → սպիտակուցային սխեմային: Սա չի փոխում հարցի էությունը, քանի որ այստեղ որոշիչ օղակը նույնպես նուկլեինաթթունն է։ Սպիտակուցից մինչև նուկլեինաթթու, ԴՆԹ կամ ՌՆԹ որոշման հակառակ ուղիներն անհայտ են:
Որպեսզի շարունակենք բջիջների կառուցվածքների ուսումնասիրությունը, որոնք կապված են սպիտակուցի սինթեզի բոլոր փուլերի հետ, մենք պետք է համառոտ կանգ առնենք այս երևույթը որոշող հիմնական գործընթացների և բաղադրիչների վրա:
Ներկայումս հիմնված ժամանակակից գաղափարներսպիտակուցների կենսասինթեզի մասին կարող ենք տալ այս բարդ և բազմաստիճան գործընթացի հետևյալ ընդհանուր սխեմատիկ դիագրամը (նկ. 16).
Սպիտակուցների սպեցիֆիկ կառուցվածքի որոշման գլխավոր, «հրամանատար» դերը պատկանում է դեզօքսիռիբոնուկլեինաթթուն՝ ԴՆԹ-ին։ ԴՆԹ-ի մոլեկուլը չափազանց երկար գծային կառուցվածք է, որը բաղկացած է երկու միահյուսված պոլիմերային շղթաներից: Այս շղթաների բաղկացուցիչ տարրերը՝ մոնոմերները, չորս տեսակի դեզօքսիռիբոնուկլեոտիդներ են, որոնց հերթափոխը կամ հաջորդականությունը շղթայի երկայնքով եզակի և հատուկ է ԴՆԹ-ի յուրաքանչյուր մոլեկուլի և նրա յուրաքանչյուր հատվածի համար։ ԴՆԹ-ի մոլեկուլի տարբեր բավական երկար հատվածներ պատասխանատու են տարբեր սպիտակուցների սինթեզի համար: Այսպիսով, մեկ ԴՆԹ մոլեկուլ կարող է որոշել սինթեզը մեծ թվովֆունկցիոնալ և քիմիապես տարբեր բջջային սպիտակուցներ: Յուրաքանչյուր տեսակի սպիտակուցի սինթեզի համար պատասխանատու է ԴՆԹ-ի մոլեկուլի միայն որոշակի հատվածը: ԴՆԹ-ի մոլեկուլի նման շրջանը, որը կապված է բջջում մեկ կոնկրետ սպիտակուցի սինթեզի հետ, հաճախ կոչվում է «ցիստրոն» տերմինով: Ներկայումս ցիստրոն հասկացությունը համարվում է գեն հասկացությանը համարժեք: Գենի եզակի կառուցվածքը՝ շղթայի երկայնքով նրա նուկլեոտիդների որոշակի հաջորդական դասավորության մեջ, պարունակում է բոլոր տեղեկությունները մեկ համապատասխան սպիտակուցի կառուցվածքի մասին։
Սկսած ընդհանուր սխեմանսպիտակուցի սինթեզ, կարելի է տեսնել (տես նկ. 16), որ ելակետը, որտեղից սկսվում է բջջում սպիտակուցների կենսասինթեզի համար տեղեկատվության հոսքը, ԴՆԹ-ն է։ Հետևաբար, հենց ԴՆԹ-ն է պարունակում տեղեկատվության առաջնային գրառումը, որը պետք է պահպանվի և վերարտադրվի բջիջից բջիջ, սերնդից սերունդ:
Հակիրճ անդրադառնալով պահպանման խնդրին գենետիկ տեղեկատվություն, այսինքն. բջջում ԴՆԹ-ի տեղայնացման մասին կարելի է ասել հետեւյալը. Վաղուց հայտնի է, որ, ի տարբերություն սպիտակուցային սինթեզող ապարատի բոլոր մյուս բաղադրիչների, ԴՆԹ-ն ունի հատուկ, շատ սահմանափակ տեղայնացում. նրա գտնվելու վայրը բարձրագույն (էուկարիոտ) օրգանիզմների բջիջներում կլինի բջջային միջուկը: Ստորին (պրոկարիոտ) օրգանիզմներում, որոնք չունեն լավ ձևավորված բջջային միջուկ, ԴՆԹ-ն նույնպես խառնվում է պրոտոպլազմայի մնացած մասից՝ մեկ կամ մի քանի կոմպակտ նուկլեոտիդային գոյացությունների տեսքով։ Դրան լիովին համապատասխան՝ էուկարիոտների միջուկը կամ պրոկարիոտների նուկլեոիդը վաղուց համարվում էր որպես գեների անոթ՝ որպես եզակի բջջային օրգանել, որը վերահսկում է օրգանիզմների ժառանգական հատկությունների իրականացումը և դրանց փոխանցումը սերունդներին:
ԴՆԹ-ի մակրոմոլեկուլային կառուցվածքի հիմքում ընկած հիմնական սկզբունքը, այսպես կոչված, փոխլրացման սկզբունքն է (նկ. 17): Ինչպես արդեն նշվեց, ԴՆԹ-ի մոլեկուլը բաղկացած է երկու միահյուսված շղթաներից։ Այս շղթաները միմյանց հետ կապված են իրենց հակադիր նուկլեոտիդների փոխազդեցության միջոցով։ Միևնույն ժամանակ, կառուցվածքային պատճառներով, նման երկշղթա կառուցվածքի առկայությունը հնարավոր է միայն այն դեպքում, եթե երկու շղթաների հակառակ նուկլեոտիդները ստերիկորեն փոխլրացնող են, այսինքն. կլինի նրանց տարածական կառուցվածքըլրացնում են միմյանց. Նման փոխլրացնող-կոմպլեմենտար զույգ նուկլեոտիդներ են զույգ Ա-Թ(ադենին-տիմին) և մի զույգ G-C (գուանին-ցիտոզին):
Հետևաբար, փոխլրացման այս սկզբունքի համաձայն, եթե ԴՆԹ-ի մոլեկուլի մեկ շղթայում ունենք չորս տեսակի նուկլեոտիդների որոշակի հաջորդականություն, ապա երկրորդ շղթայում նուկլեոտիդների հաջորդականությունը եզակիորեն կորոշվի, այնպես որ առաջին շղթայի յուրաքանչյուր Ա. Երկրորդ շղթայում կհամապատասխանի T-ին, առաջին շղթայի յուրաքանչյուր T-ին` A-ին երկրորդ շղթայում, առաջին շղթայի յուրաքանչյուր G-ին` C-ին երկրորդ շղթայում և առաջին շղթայի յուրաքանչյուր C-ին` G-ին երկրորդ շղթայում:
ԴՆԹ-ի մոլեկուլի երկշղթա կառուցվածքի հիմքում ընկած կառուցվածքային այս սկզբունքը հեշտացնում է սկզբնական կառուցվածքի ճշգրիտ վերարտադրումը, այսինքն. մոլեկուլի շղթաներում գրանցված տեղեկատվության ճշգրիտ վերարտադրումը չորս տեսակի նուկլեոտիդների հատուկ հաջորդականության տեսքով: Իրոք, բջիջում ԴՆԹ-ի նոր մոլեկուլների սինթեզը տեղի է ունենում միայն գոյություն ունեցող ԴՆԹ մոլեկուլների հիման վրա: Այս դեպքում սկզբնական ԴՆԹ-ի մոլեկուլի երկու շղթաները սկսում են շեղվել ծայրերից մեկից, և առանձնացված միաշղթա հատվածներից յուրաքանչյուրի վրա երկրորդ շղթան սկսում է հավաքվել միջավայրում առկա ազատ նուկլեոտիդներից՝ խիստ համապատասխան: փոխլրացման սկզբունքը։ ԴՆԹ-ի սկզբնական մոլեկուլի երկու շղթաների շեղման գործընթացը շարունակվում է, և համապատասխանաբար երկու շղթաներն էլ լրացվում են փոխլրացնող շղթաներով: Արդյունքում (ինչպես երևում է նկ. 17-ում), մեկի փոխարեն հայտնվում են երկու ԴՆԹ մոլեկուլներ, որոնք ճիշտ նույնական են բնօրինակին: Ստացված յուրաքանչյուր «դուստր» ԴՆԹ-ի մոլեկուլում մի շարանը ամբողջությամբ բխում է բնօրինակից, իսկ մյուսը՝ նոր սինթեզված:
Պետք է ընդգծել, որ ճշգրիտ վերարտադրության ներուժը բնորոշ է որպես այդպիսին ԴՆԹ-ի երկշղթա կոմպլեմենտար կառուցվածքին, և դրա բացահայտումը, իհարկե, կենսաբանության գլխավոր ձեռքբերումներից է:
Այնուամենայնիվ, ԴՆԹ-ի վերարտադրման (կրկնօրինակման) խնդիրը չի սահմանափակվում նրա կառուցվածքի պոտենցիալ ունակության մասին՝ իր սեփականը ճշգրիտ վերարտադրելու համար: նուկլեոտիդային հաջորդականություն. Փաստն այն է, որ ԴՆԹ-ն ինքնին ամենևին էլ ինքնավերարտադրվող մոլեկուլ չէ։ Սինթեզի գործընթացի իրականացման համար՝ ԴՆԹ-ի վերարտադրությունը վերը նկարագրված սխեմայի համաձայն, անհրաժեշտ է ԴՆԹ պոլիմերազ կոչվող հատուկ ֆերմենտային համալիրի ակտիվությունը: Հենց այս ֆերմենտն է, որ հաջորդաբար անցնում է ԴՆԹ-ի մոլեկուլի մի ծայրից մյուսը՝ երկու շղթաների բաժանման պրոցեսը դրանց վրա ազատ նուկլեոտիդների միաժամանակյա պոլիմերացումով՝ ըստ կոմպլեմենտար սկզբունքի։ Այսպիսով, ԴՆԹ-ն, ինչպես մատրիցը, սահմանում է միայն նուկլեոտիդների կարգը սինթեզված շղթաներում, իսկ գործընթացն ինքնին իրականացվում է սպիտակուցի միջոցով։ ԴՆԹ-ի վերարտադրության ժամանակ ֆերմենտի աշխատանքը ներկայումս ամենահետաքրքիր խնդիրներից է։ Հավանական է, որ ԴՆԹ պոլիմերազը ակտիվորեն սողում է երկշղթա ԴՆԹ-ի մոլեկուլի երկայնքով մի ծայրից մյուսը՝ թողնելով պատառաքաղված կրկնօրինակված «պոչը» դրա հետևում։ Այս սպիտակուցի նման աշխատանքի ֆիզիկական սկզբունքները դեռ պարզ չեն։
Այնուամենայնիվ, ԴՆԹ-ն և նրա առանձին ֆունկցիոնալ շրջանները, որոնք կրում են տեղեկատվություն սպիտակուցների կառուցվածքի մասին, իրենք ուղղակիորեն չեն մասնակցում սպիտակուցի մոլեկուլների ստեղծման գործընթացին։ ԴՆԹ-ի շղթաներում գրանցված այս տեղեկատվության իրացման առաջին քայլը այսպես կոչված տրանսկրիպցիայի կամ «վերագրման» գործընթացն է: Այս գործընթացում ԴՆԹ-ի մեկ շղթայի վրա, ինչպես մատրիցայի վրա, սինթեզվում է քիմիապես կապված պոլիմեր՝ ռիբոնուկլեինաթթու (ՌՆԹ): ՌՆԹ-ի մոլեկուլը մեկ շղթա է, որի մոնոմերները չորս տեսակի ռիբոնուկլեոտիդներ են, որոնք համարվում են ԴՆԹ-ի չորս տեսակի դեզօքսիռիբոնուկլեոտիդների աննշան ձևափոխում։ Ստացված ՌՆԹ շղթայում չորս տեսակի ռիբոնուկլեոտիդների դասավորության հաջորդականությունը ճշգրտորեն կրկնում է ԴՆԹ-ի երկու շղթաներից մեկի համապատասխան դեզօքսիռիբոնուկլեոտիդների դասավորության հաջորդականությունը։ Այս կերպ գեների նուկլեոտիդային հաջորդականությունը պատճենվում է ՌՆԹ մոլեկուլների տեսքով, այսինքն. տվյալ գենի կառուցվածքում գրանցված տեղեկատվությունը ամբողջությամբ պատճենվում է ՌՆԹ-ին: Նման «պատճենների»՝ ՌՆԹ-ի մոլեկուլների մեծ, տեսականորեն անսահմանափակ թվով կարելի է հեռացնել յուրաքանչյուր գենից։ Այս մոլեկուլները, որոնք վերագրվում են բազմաթիվ օրինակներում որպես գեների «պատճեններ» և, հետևաբար, կրում են նույն տեղեկությունը, ինչ գեները, ցրվում են ամբողջ բջջում։ Նրանք արդեն անմիջականորեն հաղորդակցվում են բջջի սպիտակուց սինթեզող մասնիկների հետ և «անձնական» մասնակցություն ունենում սպիտակուցի մոլեկուլների ստեղծման գործընթացներում։ Այսինքն՝ ինֆորմացիան այն պահված վայրից փոխանցում են այն իրացման վայրերը։ Համապատասխանաբար, այս ՌՆԹ-ները կոչվում են սուրհանդակ (mRNA) կամ սուրհանդակ (mRNA):
Պարզվել է, որ mRNA շղթան սինթեզվում է ուղղակիորեն՝ օգտագործելով համապատասխան ԴՆԹ շրջանը՝ որպես ձևանմուշ։ Սինթեզված mRNA շղթան ճշգրտորեն պատճենում է իր նուկլեոտիդային հաջորդականության երկու ԴՆԹ շղթաներից մեկը (ենթադրելով, որ ՌՆԹ-ում ուրացիլը (U) համապատասխանում է իր ածանցյալ թիմինին (T) ԴՆԹ-ում): Սա տեղի է ունենում փոխլրացման նույն կառուցվածքային սկզբունքի հիման վրա, որը որոշում է ԴՆԹ-ի կրկնօրինակումը (նկ. 18): Պարզվել է, որ երբ mRNA-ն սինթեզվում է բջջի ԴՆԹ-ի վրա, ԴՆԹ-ի միայն մեկ շղթան օգտագործվում է որպես կաղապար՝ mRNA շղթայի ձևավորման համար։ Այնուհետև այս ԴՆԹ շղթայի յուրաքանչյուր G-ն կհամապատասխանի C-ին կառուցվող ՌՆԹ-ի շղթայում, ԴՆԹ-ի յուրաքանչյուր C-ին` G-ին ՌՆԹ-ի շղթայում, ԴՆԹ-ի յուրաքանչյուր T-ին` A-ին ՌՆԹ-ի շղթայում և յուրաքանչյուր A-ին ԴՆԹ-ի շղթայում: - Y ՌՆԹ շղթայում: Արդյունքում ստացված ՌՆԹ-ի շղթան խստորեն լրացնում է ԴՆԹ-ի կաղապարի շարանը և, հետևաբար, նույնական է նուկլեոտիդային հաջորդականությամբ (ենթադրելով T=Y) ԴՆԹ-ի երկրորդ շղթային: Այսպիսով, տեղեկատվությունը «վերագրվում» է ԴՆԹ-ից ՌՆԹ, այսինքն. արտագրում. ՌՆԹ-ի շղթայում նուկլեոտիդների «վերագրված» համակցություններն արդեն ուղղակիորեն որոշում են սպիտակուցային շղթայում նրանց կողմից կոդավորված համապատասխան ամինաթթուների դասավորությունը։
Այստեղ, ինչպես ԴՆԹ-ի կրկնօրինակման դեպքում, տրանսկրիպցիոն գործընթացի ամենակարևոր ասպեկտներից մեկը պետք է լինի դրա ֆերմենտային բնույթը: ԴՆԹ-ն, որն այս գործընթացի ձևանմուշն է, ամբողջությամբ որոշում է նուկլեոտիդների դասավորությունը սինթեզված mRNA շղթայում, ստացված ՌՆԹ-ի ողջ յուրահատկությունը, բայց գործընթացն ինքնին իրականացվում է հատուկ սպիտակուցի՝ ֆերմենտի կողմից: Այս ֆերմենտը կոչվում է ՌՆԹ պոլիմերազ: Դրա մոլեկուլն ունի բարդ կազմակերպվածություն, որը թույլ է տալիս ակտիվորեն շարժվել ԴՆԹ-ի մոլեկուլի երկայնքով՝ միաժամանակ սինթեզելով ԴՆԹ-ի շղթաներից մեկին լրացնող ՌՆԹ շղթա: ԴՆԹ-ի մոլեկուլը, որը ծառայում է որպես մատրիցա, չի սպառվում և չի փոխվում՝ մնալով իր սկզբնական տեսքով և միշտ պատրաստ է նրանից անսահմանափակ թվով «պատճենների»՝ mRNA-ի նման վերագրման: Այս mRNA-ների հոսքը ԴՆԹ-ից դեպի ռիբոսոմներ կազմում է տեղեկատվության հոսք, որն ապահովում է բջջի սպիտակուցային սինթեզող ապարատի, նրա ռիբոսոմների ամբողջության ծրագրավորումը։
Այսպիսով, սխեմայի դիտարկվող մասը նկարագրում է տեղեկատվության հոսքը ԴՆԹ-ից mRNA մոլեկուլների տեսքով դեպի սպիտակուցներ սինթեզող ներբջջային մասնիկներ։ Այժմ մենք դիմում ենք այլ տեսակի հոսքի` նյութի հոսքին, որից պետք է ստեղծվի սպիտակուցը: Սպիտակուցի մոլեկուլի տարրական միավորները՝ մոնոմերները, ամինաթթուներն են, որոնցից մոտ 20-ը: Սպիտակուցի մոլեկուլ ստեղծելու (սինթեզի) համար բջջում առկա ազատ ամինաթթուները պետք է ներգրավվեն համապատասխան հոսքի մեջ, որը մտնում է սպիտակուցի սինթեզ: մասնիկ, և արդեն այնտեղ դրանք դասավորված են շղթայի մեջ որոշակի յուրահատուկ ձևով, որը թելադրված է սուրհանդակ ՌՆԹ-ով: Ամինաթթուների այս ներգրավվածությունը շինանյութսպիտակուց ստեղծելու համար - իրականացվում է ազատ ամինաթթուներ կցելով համեմատաբար փոքր չափի հատուկ ՌՆԹ մոլեկուլներին: Այս ՌՆԹ-ները, որոնք ծառայում են դրանց վրա ազատ ամինաթթուներ կցելու համար, սակայն տեղեկատվական չլինելով, ունեն այլ՝ ադապտեր, ֆունկցիա, որի իմաստը կպարզվի ավելի ուշ։ Ամինաթթուները կցվում են փոխանցման ՌՆԹ-ի (tRNA) փոքր շղթաների մի ծայրին, մեկ ամինաթթու մեկ ՌՆԹ-ի մոլեկուլին: Բջջի յուրաքանչյուր նման ամինաթթվի համար կան հատուկ ադապտեր ՌՆԹ մոլեկուլներ, որոնք միացնում են միայն այս ամինաթթուները: ՌՆԹ-ից կախված նման ձևով ամինաթթուները մտնում են սպիտակուցներ սինթեզող մասնիկներ։
Սպիտակուցների կենսասինթեզի գործընթացի կենտրոնական պահը այս երկու ներբջջային հոսքերի՝ տեղեկատվության հոսքի և նյութի հոսքի միաձուլումն է բջջի սպիտակուցային սինթեզող մասնիկների մեջ։ Այս մասնիկները կոչվում են ռիբոսոմներ: Ռիբոսոմները մոլեկուլային չափերի ուլտրամիկրոսկոպիկ կենսաքիմիական «մեքենաներ» են, որտեղ հատուկ սպիտակուցներ հավաքվում են մուտքային ամինաթթուների մնացորդներից՝ ըստ հաղորդագրության ՌՆԹ-ում պարունակվող պլանի: Թեև նկ. 19 ցույց է տալիս միայն մեկ մասնիկ, յուրաքանչյուր բջիջ կպարունակի հազարավոր կողոսկրեր: Ռիբոսոմների թիվը որոշում է բջջում սպիտակուցի սինթեզի ընդհանուր ինտենսիվությունը: Ռիբոսոմային մեկ մասնիկի տրամագիծը մոտ 20 նմ է։ Իր ձևով քիմիական բնույթՌիբոսոմ - ռիբոնուկլեոպրոտեին. այն բաղկացած է հատուկ ռիբոսոմային ՌՆԹ-ից (սա ՌՆԹ-ի երրորդ դասն է, որը մեզ հայտնի է բացի տեղեկատվության և ադապտեր ՌՆԹ-ից) և կառուցվածքային ռիբոսոմային սպիտակուցի մոլեկուլներից: Միասին մի քանի տասնյակ մակրոմոլեկուլների այս համադրությունը կազմում է իդեալական կազմակերպված և հուսալի «մեքենա», որն ունի mRNA շղթայում պարունակվող տեղեկատվությունը կարդալու և այն որոշակի կառուցվածքի պատրաստի սպիտակուցի մոլեկուլի տեսքով կարդալու ունակություն: Քանի որ գործընթացի էությունն այն է, որ սպիտակուցային շղթայում 20 տարբեր ամինաթթուների գծային դասավորությունը եզակիորեն որոշվում է չորս տարբեր նուկլեոտիդների դասավորությամբ քիմիապես բոլորովին այլ պոլիմերի՝ նուկլեինաթթվի (mRNA) շղթայում, այս գործընթացը, որը. տեղի է ունենում ռիբոսոմում, սովորաբար կոչվում է «թարգմանություն» կամ «թարգմանություն»՝ թարգմանություն, ասես, նուկլեինաթթուների շղթաների չորս տառանոց այբուբենից դեպի սպիտակուցային (պոլիպեպտիդ) շղթաների քսան տառանոց այբուբեն: Ինչպես տեսնում եք, երեքն էլ ներգրավված են թարգմանության գործընթացում: հայտնի դասՌՆԹ՝ սուրհանդակ ՌՆԹ, որը թարգմանության առարկա է. ռիբոսոմային ՌՆԹ, որը կատարում է սպիտակուցային սինթեզող ռիբոնուկլեոպրոտեին մասնիկի՝ ռիբոսոմի կազմակերպչի դերը. և ադապտեր ՌՆԹ-ներ, որոնք կատարում են թարգմանիչի ֆունկցիա:
|
Բրինձ. 19. Գործող ռիբոսոմի սխեման |

Սպիտակուցների սինթեզի գործընթացը սկսվում է ամինաթթուների միացությունների ձևավորմամբ՝ ադապտեր ՌՆԹ մոլեկուլներով կամ tRNA-ներով։ Այս դեպքում, նախ, ամինաթթվի էներգիայի «ակտիվացումը» տեղի է ունենում ադենոզին տրիֆոսֆատի (ATP) մոլեկուլի հետ նրա ֆերմենտային ռեակցիայի շնորհիվ, այնուհետև «ակտիվացված» ամինաթթուն միացված է համեմատաբար կարճ tRNA շղթայի ծայրին. մինչդեռ ակտիվացված ամինաթթվի քիմիական էներգիայի աճը պահվում է էներգիայի տեսքով քիմիական կապամինաթթուների և tRNA-ի միջև:
Միաժամանակ լուծված է երկրորդ խնդիրը. Փաստն այն է, որ ամինաթթվի և tRNA մոլեկուլի միջև ռեակցիան իրականացվում է ֆերմենտի միջոցով, որը կոչվում է ամինոացիլ-tRNA սինթետազ: 20 ամինաթթուներից յուրաքանչյուրն ունի իր հատուկ ֆերմենտները, որոնք ռեակցիան իրականացնում են միայն այս ամինաթթվի մասնակցությամբ։ Այսպիսով, կան առնվազն 20 ֆերմենտներ (aminoacyl-tRNA synthetase), որոնցից յուրաքանչյուրը հատուկ է մեկ կոնկրետ ամինաթթվի համար: Այս ֆերմենտներից յուրաքանչյուրը կարող է արձագանքել ոչ թե ցանկացած tRNA մոլեկուլի, այլ միայն նրանց հետ, որոնք իրենց շղթայում կրում են նուկլեոտիդների խիստ սահմանված համակցություն: Այսպիսով, նման հատուկ ֆերմենտների մի շարքի առկայության պատճառով, որոնք մի կողմից տարբերակում են ամինաթթվի բնույթը և, մյուս կողմից, tRNA-ի նուկլեոտիդային հաջորդականությունը, 20 ամինաթթուներից յուրաքանչյուրը «նշանակվում է»: միայն որոշակի նուկլեոտիդային համակցությամբ որոշակի tRNA-ներին:
Սխեմատիկորեն, սպիտակուցների կենսասինթեզի գործընթացի որոշ պահեր, որքանով ներկայացնում ենք դրանք այսօր, տրված են Նկ. 19. Այստեղ նախ երեւում է, որ սուրհանդակ ՌՆԹ-ի մոլեկուլը կապված է ռիբոսոմին, կամ, ինչպես ասում են, ռիբոսոմը «ծրագրավորվում» է սուրհանդակ ՌՆԹ-ի կողմից։ Ցանկացած պահի mRNA շղթայի միայն համեմատաբար կարճ հատվածն է գտնվում անմիջապես բուն ռիբոսոմում: Բայց հենց այս հատվածը՝ ռիբոսոմի մասնակցությամբ, կարող է փոխազդել ադապտեր ՌՆԹ մոլեկուլների հետ։ Այստեղ կրկին մեծ դեր է խաղում փոխլրացման սկզբունքը։
Սա այն մեխանիզմի բացատրությունն է, թե ինչու է mRNA շղթայի տրված եռյակը համապատասխանում խիստ սահմանված ամինաթթվի: Ադապտոր ՌՆԹ-ն (tRNA) անհրաժեշտ միջանկյալ օղակ է կամ ադապտեր, երբ յուրաքանչյուր ամինաթթու «ճանաչում է» իր եռյակը mRNA-ի վրա:
Նկ. Նկար 19-ը ցույց է տալիս, որ բացի tRNA մոլեկուլից, կցված ամինաթթուով, ռիբոսոմում կա ևս մեկ tRNA մոլեկուլ: Բայց, ի տարբերություն վերը քննարկված tRNA մոլեկուլի, այս tRNA մոլեկուլն իր ծայրով կցված է սինթեզի գործընթացում գտնվող սպիտակուցի (պոլիպեպտիդ) շղթայի ծայրին: Այս դիրքն արտացոլում է սպիտակուցի մոլեկուլի սինթեզի ժամանակ ռիբոսոմում տեղի ունեցող իրադարձությունների դինամիկան։ Այս դինամիկան կարելի է պատկերացնել հետևյալ կերպ. Սկսենք որոշ միջանկյալ կետից, որը ցույց է տրված Նկ. 19 և բնութագրվում է սպիտակուցային շղթայի առկայությամբ, որն արդեն սկսել է կառուցվել, դրան կցված tRNA և որը նոր է մտել ռիբոսոմ և կապված է նոր tRNA մոլեկուլի եռյակի հետ՝ իր համապատասխան ամինաթթուով: Ըստ երևույթին, tRNA մոլեկուլը ռիբոսոմի տվյալ վայրում տեղակայված mRNA եռյակին միացնելու գործողությունը հանգեցնում է այնպիսի փոխադարձ կողմնորոշման և սերտ շփման ամինաթթուների մնացորդի և կառուցվող սպիտակուցային շղթայի միջև, կովալենտային կապ. Կապը տեղի է ունենում այնպես, որ կառուցվող սպիտակուցային շղթայի վերջը (կցված է tRNA-ին նկ. 19-ում) այս tRNA-ից տեղափոխվում է մուտքային aminoacyl-tRNA-ի ամինաթթվի մնացորդ: Արդյունքում «ճիշտ» tRNA-ն, խաղալով «դոնորի» դերը, կլինի ազատ, և սպիտակուցային շղթա- փոխանցվել է «ընդունողին», այսինքն. «ձախ» (մուտքային) aminoacyl-tRNA-ի վրա: Արդյունքում սպիտակուցային շղթան կընդլայնվի մեկ ամինաթթուով և կկցվի «ձախ» tRNA-ին։ Դրան հաջորդում է «ձախ» tRNA-ն՝ դրա հետ կապված mRNA նուկլեոտիդների եռյակի հետ միասին դեպի աջ, այնուհետև նախկին «դոնոր» tRNA մոլեկուլը կտեղափոխվի այստեղից և կլքի ռիբոսոմները: Իր տեղում կհայտնվի նոր tRNA-ն՝ կառուցվող սպիտակուցային շղթայով, որը երկարաձգվում է մեկ ամինաթթվի մնացորդով, և mRNA շղթան ռիբոսոմից մեկ եռակի դեպի աջ կշարժվի: mRNA շղթան մեկ եռյակը դեպի աջ տեղափոխելու արդյունքում հաջորդ դատարկ եռյակը (UUU) կհայտնվի ռիբոսոմում, իսկ համապատասխան tRNA-ն ամինաթթուով (ֆենիլալանիլ-tRNA) կոմպլեմենտար սկզբունքով անմիջապես կմիանա դրան։ Սա կրկին կհանգեցնի կովալենտային (պեպտիդային) կապի ձևավորմանը կառուցվող սպիտակուցային շղթայի և ֆենիլալանինի մնացորդի միջև, և դրանից հետո mRNA շղթան մեկ եռյակ կտեղափոխվի աջ՝ դրանից բխող բոլոր հետևանքներով և այլն։ Այս կերպ, հաջորդաբար, եռյակ առ եռյակ, տեղեկատվական ՌՆԹ-ի շղթան ձգվում է ռիբոսոմի միջով, ինչի արդյունքում մՌՆԹ շղթան «կարդում» է ռիբոսոմն ամբողջությամբ՝ սկզբից մինչև վերջ։ Միաժամանակ և դրա հետ մեկտեղ տեղի է ունենում սպիտակուցային շղթայի հաջորդական ամինաթթու կուտակում ամինաթթվի կողմից: Համապատասխանաբար, ամինաթթուներով tRNA մոլեկուլները մեկը մյուսի հետևից մտնում են ռիբոսոմ, իսկ tRNA մոլեկուլները առանց ամինաթթուների դուրս են գալիս: Գտնվելով ռիբոսոմից դուրս լուծույթում՝ ազատ tRNA մոլեկուլները կրկին միավորվում են ամինաթթուների հետ և նորից տեղափոխում դրանք ռիբոսոմի մեջ, իրենք՝ այդպիսով, ցիկլային շրջանառության մեջ՝ առանց ոչնչացման և փոփոխության:
Կենսաբանական հաջորդականություններում պարունակվող տեղեկատվությունը
Կենսապոլիմերները (կենսաբանական) պոլիմերներ են, որոնք սինթեզվում են կենդանի էակների կողմից։ ԴՆԹ-ն, ՌՆԹ-ն և սպիտակուցները գծային պոլիմերներ են, այսինքն՝ դրանց պարունակած յուրաքանչյուր մոնոմեր միավորվում է առնվազն երկու այլ մոնոմերի հետ։ Մոնոմերների հաջորդականությունը կոդավորում է տեղեկատվություն, որի փոխանցման կանոնները նկարագրված են կենտրոնական դոգմայով։ Տեղեկատվությունը փոխանցվում է բարձր ճշգրտությամբ, դետերմինիստականորեն, և մեկ բիոպոլիմեր օգտագործվում է որպես ձևանմուշ մեկ այլ պոլիմերի հավաքման համար այն հաջորդականությամբ, որն ամբողջությամբ որոշվում է առաջին պոլիմերի հաջորդականությամբ:
Կենսաբանական տեղեկատվության փոխանցման ունիվերսալ ուղիներ
Կենդանի օրգանիզմներում կան երեք տեսակի տարասեռ, այսինքն՝ բաղկացած տարբեր պոլիմերային մոնոմերներից՝ ԴՆԹ, ՌՆԹ և սպիտակուց։ Նրանց միջև տեղեկատվության փոխանցումը կարող է իրականացվել 3 × 3 = 9 եղանակով: Կենտրոնական դոգման տեղեկատվության փոխանցման այս 9 տեսակները բաժանում է երեք խմբի.
- Ընդհանուր - հայտնաբերվել է կենդանի օրգանիզմների մեծ մասում;
- Հատուկ - հայտնաբերվել է որպես բացառություն վիրուսների և գենոմի շարժական տարրերի կամ կենսաբանական փորձի պայմաններում.
- Անհայտ - չի գտնվել:
ԴՆԹ-ի վերարտադրություն (ԴՆԹ → ԴՆԹ)
ԴՆԹ-ն կենդանի օրգանիզմների սերունդների միջև տեղեկատվության փոխանցման հիմնական միջոցն է, ուստի ԴՆԹ-ի ճշգրիտ կրկնօրինակումը (կրկնօրինակումը) շատ կարևոր է: Կրկնօրինակումն իրականացվում է սպիտակուցների համալիրի միջոցով, որը քամում է քրոմատինը, այնուհետև կրկնակի պարույրը։ Դրանից հետո ԴՆԹ պոլիմերազը և դրա հետ կապված սպիտակուցները երկու շղթաներից յուրաքանչյուրի վրա կառուցում են նույնական պատճեն:
Տրանսկրիպցիա (ԴՆԹ → ՌՆԹ)
Տրանսկրիպցիան կենսաբանական գործընթաց է, որի արդյունքում ԴՆԹ հատվածում պարունակվող տեղեկատվությունը պատճենվում է սինթեզված mRNA մոլեկուլի վրա։ Տրանսկրիպցիան իրականացվում է տրանսկրիպցիոն գործոնների և ՌՆԹ պոլիմերազի միջոցով։ Էուկարիոտիկ բջիջում առաջնային տառադարձությունը (նախա-ՌՆԹ) հաճախ խմբագրվում է: Այս գործընթացը կոչվում է միացում:
Գենետիկական տեղեկատվության իրացման սխեմատիկ դիագրամ պրո- և էուկարիոտներում:ՊՐՈԿԱՐԻՈՏՆԵՐ. Պրոկարիոտներում սպիտակուցի սինթեզը ռիբոսոմի կողմից (թարգմանություն) տարածականորեն առանձնացված չէ տրանսկրիպցիայից և կարող է տեղի ունենալ նույնիսկ մինչև ՌՆԹ պոլիմերազի կողմից mRNA սինթեզի ավարտը։ Պրոկարիոտիկ mRNA-ները հաճախ պոլիցիստրոնիկ են, այսինքն՝ պարունակում են մի քանի անկախ գեն:
ԵՎԿԱՐԻՈՏՆԵՐ. էուկարիոտիկ mRNA-ն սինթեզվում է որպես պրեկուրսոր՝ նախամՌՆԹ, որն այնուհետև ենթարկվում է բարդ փուլային հասունացման՝ վերամշակման, ներառյալ գլխարկի կառուցվածքի կցումը մոլեկուլի 5 «վերջին, մի քանի տասնյակ ադենինի մնացորդների կցումը նրա 3» ծայրին։ (պոլիադենիլացիա), աննշան հատվածների ճեղքումը՝ ինտրոններ և զգալի հատվածների միմյանց հետ կապը՝ էկզոններ (սպլացինգ)։ Այս դեպքում կարող է տեղի ունենալ նույն նախամՌՆԹ-ի էկզոնների միացումը տարբեր ճանապարհներ, որը հանգեցնում է տարբեր հասուն mRNA-ների և, ի վերջո, տարբեր սպիտակուցային տարբերակների ձևավորմանը (այլընտրանքային միացում): Միայն հաջողությամբ մշակված mRNA-ն արտահանվում է միջուկից ցիտոպլազմա և ներգրավվում թարգմանության մեջ:
Թարգմանություն (ՌՆԹ → սպիտակուց)
ՌՆԹ-ի վերարտադրություն (ՌՆԹ → ՌՆԹ)
ՌՆԹ-ի վերարտադրություն - ՌՆԹ-ի շղթայի պատճենում իր լրացուցիչ ՌՆԹ շղթային՝ օգտագործելով ՌՆԹ-ից կախված ՌՆԹ պոլիմերազ ֆերմենտը: Միաշղթա պարունակող վիրուսները (օրինակ՝ պիկորնավիրուսները, որոնք ներառում են ոտքի և բերանի հիվանդության վիրուսը) կամ կրկնակի շղթա ՌՆԹ կրկնօրինակվում են նույն ձևով։
Սպիտակուցի ուղղակի թարգմանություն ԴՆԹ կաղապարի վրա (ԴՆԹ → սպիտակուց)
Կենդանի թարգմանությունը ցուցադրվել է E. coli բջիջների քաղվածքներում, որոնք պարունակում էին ռիբոսոմներ, բայց ոչ mRNA: Նման քաղվածքները սինթեզում էին ԴՆԹ-ից համակարգ մուտքագրված սպիտակուցներ, և հակաբիոտիկ նեոմիցինը ուժեղացնում էր այդ ազդեցությունը:
Էպիգենետիկ փոփոխություններ
Էպիգենետիկ փոփոխությունները գեների արտահայտման փոփոխություններն են, որոնք պայմանավորված չեն գենետիկական տեղեկատվության փոփոխություններով (մուտացիաներ): Էպիգենետիկ փոփոխությունները տեղի են ունենում գեների արտահայտման մակարդակի փոփոխության, այսինքն՝ դրանց տրանսկրիպացիայի և/կամ թարգմանության արդյունքում։ Էպիգենետիկ կարգավորման ամենաուսումնասիրված տեսակը ԴՆԹ մեթիլացումն է ԴՆԹ մեթիլտրանսֆերազային սպիտակուցների օգնությամբ, ինչը հանգեցնում է մեթիլացված գենի ժամանակավոր, կյանքից կախված անակտիվացմանը: Այնուամենայնիվ, քանի որ ԴՆԹ-ի մոլեկուլի առաջնային կառուցվածքը չի փոխվում, այս բացառությունը չի կարող համարվել սպիտակուցից ԴՆԹ տեղեկատվության փոխանցման իրական օրինակ:
պրիոններ
Պրիոնները սպիտակուցներ են, որոնք գոյություն ունեն երկու ձևով. Սպիտակուցի ձևերից (կոնֆորմացիաներից) մեկը ֆունկցիոնալ է, սովորաբար լուծվում է ջրում։ Երկրորդ ձևը առաջացնում է ջրում չլուծվող ագրեգատներ, հաճախ մոլեկուլային պոլիմերային խողովակների տեսքով։ Մոնոմերը՝ սպիտակուցի մոլեկուլը, այս կոնֆորմացիայի մեջ ի վիճակի է միավորվել այլ նմանատիպ սպիտակուցային մոլեկուլների հետ՝ դրանք վերածելով երկրորդ, պրիոնման կոնֆորմացիայի: Սնկերի մեջ նման մոլեկուլները կարող են ժառանգվել: Բայց, ինչպես ԴՆԹ-ի մեթիլացման դեպքում, այս դեպքում էլ սպիտակուցի առաջնային կառուցվածքը մնում է նույնը, և նուկլեինաթթուներին տեղեկատվության փոխանցում չկա։
«Դոգմա» տերմինի պատմությունը
բնօրինակ տեքստ(անգլերեն)
Իմ միտքն այն էր, որ դոգման գաղափար է, որի համար ողջամիտ ապացույց չկա: Տեսնու՞մ ես։— Եվ Կրիկը ուրախությունից մռնչաց։ «Ես ուղղակի չգիտեի, թե ինչ է նշանակում դոգմա։ Եվ ես կարող էի այն անվանել «Կենտրոնական վարկած», կամ, գիտե՞ս։ ասեմ, որ դոգման պարզապես բռնելու արտահայտություն էր
Բացի այդ, իր «What a Mad Pursuit» ինքնակենսագրական գրքում Քրիկը գրել է «դոգմա» բառի ընտրության և այդ ընտրության առաջացրած խնդիրների մասին.
«Ես այս գաղափարը անվանել եմ կենտրոնական դոգմա, կարծում եմ, երկու պատճառով. Ես արդեն օգտագործել եմ հիպոթեզ բառը հաջորդականության հիպոթեզում, բացի այդ, ուզում էի առաջարկել, որ այս նոր ենթադրությունն ավելի կենտրոնական և ուժեղ է... Ինչպես պարզվում է, դոգմա տերմինի օգտագործումն ավելի շատ դժվարություններ առաջացրեց, քան արժեր... Շատերը տարիներ անց Ժակ Մոնոդն ինձ ասաց, որ, ըստ երևույթին, ես չեմ հասկացել, թե ինչ է նշանակում դոգմա բառը, որը նշանակում է հավատքի մի մասը, որը կասկածի ենթակա չէ։ Ես անորոշ մտավախություն ունեի բառի այս իմաստի վերաբերյալ, բայց քանի որ հավատում էի, որ բոլոր կրոնական համոզմունքները հիմք չունեն, ես բառն օգտագործեցի այնպես, ինչպես հասկացա, և ոչ թե այլ մարդկանց մեծ մասը, կիրառելով այն մեծ վարկածի վրա, որ չնայած վստահությանը ոգեշնչված, հիմնված էր փոքր քանակությամբ ուղղակի փորձարարական տվյալների վրա:
բնօրինակ տեքստ(անգլերեն)
Ես այս գաղափարը անվանեցի կենտրոնական դոգմա, երկու պատճառով, ես կասկածում եմ: Ես արդեն օգտագործել էի ակնհայտ հիպոթեզ բառը հաջորդականության հիպոթեզում, և ի լրումն, ես ուզում էի ենթադրել, որ այս նոր ենթադրությունն ավելի կենտրոնական և հզոր է: ... Ինչպես պարզվեց, դոգմա բառի օգտագործումը համարյա ավելի շատ դժվարություններ առաջացրեց, քան արժեր... Շատ տարիներ անց Ժակ Մոնոդը մատնանշեց ինձ, որ ես կարծես թե չեմ հասկանում. եւԴոգմա բառի ճիշտ օգտագործումը, որը համոզմունք է, որը չի կարելի կասկածել: Ես սա հասկացա անորոշ ձևով, բայց քանի որ կարծում էի, որ բոլոր կրոնական համոզմունքներն անհիմն են, ես բառն օգտագործեցի այնպես, ինչպես ես ինքս մտածում էի դրա մասին, ոչ այնպես, ինչպես անում է աշխարհի մեծ մասը, և պարզապես այն կիրառեցի մի մեծ վարկածի վրա: որը, որքան էլ հավանական է, ուներ քիչ ուղղակի փորձարարական աջակցություն:
տես նաեւ
Նշումներ
Հղումներ
- B. J. McCarthy, J. J. Holland.Դենատուրացված ԴՆԹ-ն որպես ուղղակի ձևանմուշ արհեստական պայմաններումՍպիտակուցների սինթեզ // PNAS. - 1965. - T. 54. - S. 880-886.
- Վերները, Է.Գենոմի իմաստաբանություն, սիլիկո բազմաբջիջ համակարգերում և կենտրոնական դոգման // FEBS Նամակներ. - 2005. - V. 579. - S. 1779-1782 թթ. PMID 15763551
- Հորաս Ֆրիլենդ Ջադսոն.Գլուխ 6. Իմ միտքն այն էր, որ դոգման գաղափար է, որի համար ողջամիտ ապացույց չկա: Դու տես?! // Արարման ութերորդ օրը. Կենսաբանության մեջ հեղափոխության հեղինակները (25-րդ տարեդարձի հրատարակություն): - 1996 թ.
Մոլեկուլային կենսաբանության կենտրոնական դոգմա
Բջջի միջուկի կառուցվածքը
Բջիջների ֆրակցիոնացում Այսօր ֆրակցիոնացումը հնարավորություն է տալիս ձեռք բերել գրեթե ցանկացած բջջային օրգանելներ և կառուցվածքներ՝ միջուկներ, միջուկներ, քրոմատին, միջուկային թաղանթներ, պլազմային թաղանթ, էնդոպլազմային ցանցի վակուոլներ և այլն։
Հատուկ մեթոդներ
Մինչ բջջային ֆրակցիաներ ստանալը, բջիջները քայքայվում են համասեռացման միջոցով։ Այնուհետև, ֆրակցիաները մեկուսացված են միատարր նյութերից: Բջջային կառուցվածքների մեկուսացման հիմնական մեթոդը առանձնացնող ցենտրիֆուգումն է: Այն հիմնված է այն փաստի վրա, որ ավելի ծանր մասնիկները ավելի արագ նստում են ցենտրիֆուգային խողովակի հատակին:
Ցածր արագացումների դեպքում (1-3 հազար գ) միջուկներն ու անձեռնմխելի բջիջները նստում են ավելի վաղ՝ 15-30 հազար գ-ում, ավելի մեծ մասնիկները կամ մակրոսոմները՝ կազմված միտոքոնդրիաներից, փոքր պլաստիդներից, պերօքսիսոմներից, լիզոսոմներից և այլն, նստում են 50 հազար գ-ում։ , միկրոզոմները, բջջի վակուոլային համակարգի բեկորները նստում են։ Խառը ենթաֆրակցիաները նորից ցենտրիֆուգելիս մաքուր ֆրակցիաները մեկուսացվում են: Ֆրակցիաների ավելի նուրբ տարանջատման համար օգտագործվում է սախարոզայի խտության գրադիենտ ցենտրիֆուգացիա: Առանձին բջջային բաղադրիչների ձեռքբերումը հնարավորություն է տալիս ուսումնասիրել դրանց կենսաքիմիական և ֆունկցիոնալ առանձնահատկությունները, ստեղծել առանց բջջային համակարգերի, օրինակ,ռիբոսոմների համար, որոնք կարող են սինթեզել սպիտակուցը ըստ փորձարարի կողմից նշված սուրհանդակային ՌՆԹ-ի կամ բջջային վերմոլեկուլային կառուցվածքների վերստեղծման համար։
Տեղակայված է ref.rf
Նման արհեստական համակարգերն օգնում են ուսումնասիրել բջջում տեղի ունեցող նուրբ գործընթացները։
Մեթոդ բջջային ճարտարագիտություն. Հատուկ բուժումից հետո տարբեր կենդանի բջիջները կարող են միաձուլվել միմյանց հետ և ձևավորել երկմիջուկային բջիջ կամ հետերոկարիոն: Հետերոկարիոնները, հատկապես նրանք, որոնք ձևավորվել են սերտորեն կապված բջիջներից (օրինակ՝ մկներ և համստերներ), կարող են մտնել միտոզ և առաջացնել իսկական հիբրիդային բջիջներ։ Այլ տեխնիկան հնարավորություն է տալիս բջիջներ կառուցել տարբեր ծագման միջուկներից և ցիտոպլազմայից:
Այսօր բջջային ճարտարագիտությունը լայնորեն կիրառվում է ոչ միայն փորձարարական կենսաբանության, այլև կենսատեխնոլոգիայի մեջ։ Օրինակ՝ մոնոկլոնալ հակամարմիններ ստանալու ժամանակ։
Բջիջն ունի հսկայական քանակությամբ բազմազան գործառույթներ, այդ գործառույթների կատարման հիմնական աշխատանքային մեխանիզմները սպիտակուցներն են կամ դրանց բարդույթները այլ կենսաբանական մակրոմոլեկուլների հետ: Սինթեզի, քայքայման, տարբեր սպիտակուցների, նուկլեինաթթուների, լիպիդների, ածխաջրերի գրեթե բոլոր գործընթացները տեղի են ունենում ֆերմենտային սպիտակուցների մասնակցությամբ։ Կծկումը, որը հանգեցնում է բջիջների շարժունակությանը կամ բջիջների ներսում նյութերի և կառուցվածքների շարժմանը, իրականացվում է նաև հատուկ կծկվող սպիտակուցների միջոցով։ Բջջային շատ ռեակցիաներ՝ ի պատասխան արտաքին գործոնների (վիրուսներ, հորմոններ, օտար սպիտակուցներ և այլն) սկսվում են այդ գործոնների փոխազդեցությամբ բջջային հատուկ ընկալիչների սպիտակուցների հետ։
Սպիտակուցները գրեթե բոլոր բջջային կառույցների հիմնական բաղադրիչներն են:
Տեղակայված է ref.rf
Յուրաքանչյուր առանձին սպիտակուցի կառուցվածքը խիստ սպեցիֆիկ է, որն արտահայտվում է դրանց առաջնային կառուցվածքի յուրահատկությամբ՝ պոլիպեպտիդային, սպիտակուցային շղթայի երկայնքով ամինաթթուների հաջորդականությամբ։ Սպիտակուցային շղթայում ամինաթթուների միանշանակ հաջորդականությունը վերարտադրելու նման կոռեկտությունը որոշվում է այդ գենային շրջանի ԴՆԹ-ի կառուցվածքով, որն ի վերջո պատասխանատու է այս սպիտակուցի կառուցվածքի և սինթեզի համար: Այս դիրքորոշումը մոլեկուլային կենսաբանության հիմնական պոստուլատն է կամ նրա «դոգմա»-ն։ Բացի այդ, կենտրոնական դոգման ընդգծում է տեղեկատվության միակողմանի փոխանցումը՝ միայն ԴՆԹ-ից սպիտակուց (DNA ® mRNA ® սպիտակուց) և հերքում է վերադարձի ուղիները՝ սպիտակուցից մինչև նուկլեինաթթու:
Ներկայիս գիտելիքների հիման վրա սպիտակուցի կենսասինթեզը հետևյալ սխեմատիկ դիագրամն է.
գլխավոր դերըսպիտակուցների հատուկ կառուցվածքը որոշելիս պատկանում է ԴՆԹ-ին: ԴՆԹ-ի մոլեկուլը, որը բաղկացած է երկու խճճված պոլիմերային շղթայից, գծային կառուցվածք է, որի մոնոմերները չորս տեսակի դեզօքսիրիբոնուկլեոտիդներ են, որոնց հերթափոխը կամ հաջորդականությունը շղթայի երկայնքով եզակի է և հատուկ ԴՆԹ-ի յուրաքանչյուր մոլեկուլի և նրա յուրաքանչյուր հատվածի համար: ԴՆԹ-ի մոլեկուլի որոշակի հատվածը պատասխանատու է յուրաքանչյուր սպիտակուցի սինթեզի համար: ԴՆԹ-ի մոլեկուլի մի հատված, որը պարունակում է բոլոր տեղեկությունները մեկ համապատասխան սպիտակուցի կառուցվածքի մասին: կոչվում է ցիստրոն: Այսօր ցիստրոն հասկացությունը համարվում է գեն հասկացությանը համարժեք:
Հայտնի է, որ, ի տարբերություն սպիտակուցային սինթեզող ապարատի այլ բաղադրիչների, բջիջներում էուկարիոտիկ օրգանիզմների ԴՆԹ-ի գտնվելու վայրը բջջի միջուկն է։ Ստորին (պրոկարիոտ) օրգանիզմներում, որոնք չունեն լավ ձևավորված բջջի միջուկ, ԴՆԹ-ն առանձնացված է նաև պրոտոպլազմայի մնացած մասերից մեկ կամ մի քանի կոմպակտ նուկլեոտիդների տեսքով։
ԴՆԹ-ի մակրոմոլեկուլային կառուցվածքի հիմքում ընկած է այսպես կոչված փոխլրացման սկզբունքը։ Դա նշանակում է, որ երկու միահյուսված ԴՆԹ-ի շղթաների հակառակ նուկլեոտիդները լրացնում են միմյանց իրենց տարածական կառուցվածքով։ Նման փոխլրացնող-կոմպլեմենտար նուկլեոտիդային զույգերն են՝ A-T զույգը (ադենին-թիմին) և G-C զույգը (գուանին-ցիտոզին):
Բջջում ԴՆԹ-ի նոր մոլեկուլների սինթեզը տեղի է ունենում միայն գոյություն ունեցող ԴՆԹ մոլեկուլների հիման վրա: Այս դեպքում սկզբնական ԴՆԹ-ի մոլեկուլի երկու շղթաները սկսում են շեղվել ծայրերից մեկից, և առանձնացված միաշղթա հատվածներից յուրաքանչյուրի վրա երկրորդ շղթան սկսում է հավաքվել միջավայրում առկա ազատ նուկլեոտիդներից՝ խիստ համապատասխան: փոխլրացման սկզբունքը։ ԴՆԹ-ի յուրաքանչյուր «դուստր» մոլեկուլում մի շղթան ամբողջությամբ բխում է բնօրինակից, իսկ մյուսը՝ նոր սինթեզված:
Պետք է ընդգծել, որ ճշգրիտ վերարտադրության ներուժը բնորոշ է հենց ԴՆԹ-ի երկշղթա կոմպլեմենտար կառուցվածքին, և դրա բացահայտումը կենսաբանության հիմնական ձեռքբերումներից է:
Սինթեզի՝ ԴՆԹ-ի վերարտադրության գործընթացն իրականացնելու համար վերը նկարագրված սխեմայի համաձայն, անհրաժեշտ է ԴՆԹ պոլիմերազ կոչվող հատուկ ֆերմենտի ակտիվությունը։ Հենց այս ֆերմենտն է, որ հաջորդաբար անցնում է ԴՆԹ-ի մոլեկուլի մի ծայրից մյուսը՝ երկու շղթաների բաժանման պրոցեսը դրանց վրա ազատ նուկլեոտիդների միաժամանակյա պոլիմերացումով՝ ըստ կոմպլեմենտար սկզբունքի։
Հետևաբար, ԴՆԹ-ն, ինչպես մատրիցը, սահմանում է միայն նուկլեոտիդների կարգը սինթեզված շղթաներում, և գործընթացն ինքնին իրականացվում է սպիտակուցի միջոցով։ ԴՆԹ-ն և նրա առանձին ֆունկցիոնալ շրջանները, որոնք կրում են տեղեկատվություն սպիտակուցների կառուցվածքի մասին, իրենք ուղղակիորեն չեն մասնակցում սպիտակուցի մոլեկուլների ստեղծման գործընթացին։ Այս տեղեկատվության իրացմանն ուղղված առաջին քայլը, այսպես կոչված, արտագրման գործընթացն է կամ «վերագրելը»: Այս գործընթացում քիմիապես կապված պոլիմեր՝ ռիբոնուկլեինաթթու (ՌՆԹ), սինթեզվում է ԴՆԹ շղթայի վրա, ինչպես մատրիցայի վրա։ ՌՆԹ մոլեկուլը մեկ շղթա է, որի մոնոմերները չորս տեսակի ռիբոնուկլեոտիդներ են։ Ստացված ՌՆԹ շղթայում չորս տեսակի ռիբոնուկլեոտիդների դասավորության հաջորդականությունը ճշգրտորեն կրկնում է ԴՆԹ-ի երկու շղթաներից մեկի համապատասխան դեզօքսիռիբոնուկլեոտիդների դասավորության հաջորդականությունը։ Դրա շնորհիվ այս գենի կառուցվածքում գրանցված տեղեկատվությունը ամբողջությամբ պատճենվում է ՌՆԹ-ին։ Տեսականորեն անսահմանափակ թվով «պատճեններ»՝ ՌՆԹ մոլեկուլներ, կարող են հեռացվել յուրաքանչյուր գենից: ՌՆԹ մոլեկուլները հաղորդակցության մեջ են մտնում բջջի սպիտակուցը սինթեզող մասնիկների հետ և անմիջականորեն մասնակցում են սպիտակուցի մոլեկուլների սինթեզին։ Այսինքն՝ ինֆորմացիան դրա պահպանման վայրերից փոխանցում են իրականացման վայրեր։ Այդ իսկ պատճառով այդ ՌՆԹ-ները կոչվում են սուրհանդակ կամ սուրհանդակ ՌՆԹ, կրճատ՝ mRNA կամ mRNA:
Սինթեզված սուրհանդակային ՌՆԹ շղթան ուղղակիորեն օգտագործում է համապատասխան ԴՆԹ շրջանը որպես ձևանմուշ։ Այս դեպքում սինթեզված mRNA շղթան ճշգրիտ կերպով պատճենում է իր նուկլեոտիդային հաջորդականության երկու ԴՆԹ շղթաներից մեկը (ՌՆԹ-ում ուրացիլը (U) համապատասխանում է իր ածանցյալ թիմինին (T) ԴՆԹ-ում): Ամեն ինչ տեղի է ունենում փոխլրացման նույն սկզբունքի հիման վրա, որը որոշում է ԴՆԹ-ի կրկնօրինակումը: Արդյունքում տեղի է ունենում տեղեկատվության «վերագրում» կամ վերագրում ԴՆԹ-ից ՌՆԹ: ՌՆԹ նուկլեոտիդների «վերագրված» համակցություններն արդեն ուղղակիորեն որոշում են սպիտակուցային շղթայում նրանց կողմից կոդավորված ամինաթթուների դասավորությունը:
Հիմա ինչպե՞ս է պատրաստվում սպիտակուցը: Հայտնի է, որ սպիտակուցի մոլեկուլի մոնոմերների տեսակները ամինաթթուներն են, որոնցից կան 20 տարբեր սորտեր։ Բջջի ամինաթթվի յուրաքանչյուր տեսակի համար կան հատուկ ադապտեր ՌՆԹ մոլեկուլներ, որոնք կցում են միայն այս տեսակի ամինաթթուները: ՌՆԹ-ում այցելած ձևով ամինաթթուները մտնում են սպիտակուց սինթեզող մասնիկներ՝ ռիբոսոմներ, և արդեն այնտեղ, սուրհանդակ ՌՆԹ-ի թելադրանքով, տեղադրվում են սինթեզված սպիտակուցի շղթայում։
Սպիտակուցների կենսասինթեզի հիմնական բանը ռիբոսոմներում երկու ներբջջային հոսքերի միաձուլումն է՝ տեղեկատվության հոսքը և նյութի հոսքը: Ռիբոսոմները մոլեկուլային չափերի կենսաքիմիական «մեքենաներ» են, որոնցում հատուկ սպիտակուցներ հավաքվում են մուտքային ամինաթթուների մնացորդներից՝ ըստ հաղորդագրության ՌՆԹ-ում պարունակվող պլանի։ Յուրաքանչյուր բջիջ պարունակում է հազարավոր կողոսկրեր, սպիտակուցների սինթեզի ինտենսիվությունը որոշվում է բջիջում դրանց քանակով։ Իր քիմիական բնույթով ռիբոսոմը պատկանում է ռիբոնուկլեոպրոտեիններին և բաղկացած է հատուկ ռիբոսոմային ՌՆԹ-ից և ռիբոսոմային սպիտակուցի մոլեկուլներից։ Ռիբոսոմները կարող են կարդալ mRNA շղթայում պարունակվող տեղեկատվությունը և այն իրականացնել պատրաստի սպիտակուցի մոլեկուլի տեսքով: Գործընթացի էությունը կայանում է նրանում, որ սպիտակուցային շղթայում 20 տեսակի ամինաթթուների գծային դասավորությունը որոշվում է չորս տեսակի նուկլեոտիդների դասավորությամբ բոլորովին այլ պոլիմերի՝ նուկլեինաթթվի (mRNA) շղթայում: Այդ իսկ պատճառով, ռիբոսոմում տեղի ունեցող այս գործընթացը սովորաբար կոչվում է «թարգմանություն» կամ «թարգմանություն»՝ թարգմանություն նուկլեինաթթուների 4 տառանոց այբուբենից դեպի սպիտակուցային (պոլիպեպտիդ) շղթաների 20 տառանոց այբուբեն: Այս թարգմանության գործընթացում ներգրավված են ՌՆԹ-ի բոլոր երեք հայտնի դասերը՝ մեսենջեր ՌՆԹ, որը թարգմանության առարկա է, ռիբոսոմային ՌՆԹ, որը կատարում է ռիբոսոմի կազմակերպչի դերը և ադապտեր ՌՆԹ, որը հանդես է գալիս որպես թարգմանիչ։
Սպիտակուցների սինթեզի գործընթացը սկսվում է ամինաթթուների միացությունների առաջացմամբ՝ ադապտեր ՌՆԹ մոլեկուլներով։ Այս դեպքում, նախ, ամինաթթվի «ակտիվացում» էներգիան տեղի է ունենում ադենոզին տրիֆոսֆատի (ATP) մոլեկուլի հետ նրա ֆերմենտային ռեակցիայի շնորհիվ, այնուհետև «ակտիվացված» ամինաթթուն միանում է համեմատաբար կարճ tRNA շղթայի ծայրին, մինչդեռ ավելանում է. ակտիվացված ամինաթթվի քիմիական էներգիան պահվում է ամինաթթվի և tRNA-ի միջև քիմիական կապի էներգիայի տեսքով:
Ավելացնենք, որ ամինաթթվի և tRNA մոլեկուլի միջև ռեակցիան իրականացվում է ամինոացիլ-tRNA սինթետազ ֆերմենտի միջոցով։ 20 ամինաթթուներից յուրաքանչյուրն ունի իր ֆերմենտները, որոնք ռեակցիան իրականացնում են միայն այս ամինաթթվի մասնակցությամբ։
Մոլեկուլային կենսաբանության կենտրոնական դոգման հասկացությունն ու տեսակներն են։ «Մոլեկուլային կենսաբանության կենտրոնական դոգմա» կատեգորիայի դասակարգումը և առանձնահատկությունները 2017, 2018 թ.
Մատրիցային կենսասինթեզի հիմնական գործիչը նուկլեինաթթուներն են՝ ՌՆԹ և ԴՆԹ: Դրանք պոլիմերային մոլեկուլներ են, որոնք ներառում են հինգ տեսակի ազոտային հիմքեր, երկու տեսակի պենտոզներ և ֆոսֆորաթթվի մնացորդներ։ Նուկլեինաթթուներում ազոտային հիմքերը կարող են լինել պուրինը (ադենին, գուանին) և պիրիմիդինը (ցիտոզին, ուրացիլը (միայն ՌՆԹ-ում), թիմինը (միայն ԴՆԹ-ում)): Կախված ածխաջրերի կառուցվածքից. ռիբոնուկլեինաթթուներ - պարունակում է ռիբոզ (ՌՆԹ) և դեզօքսիռիբոնուկլեինաթթուներ- պարունակում է դեզօքսիռիբոզ (ԴՆԹ):
Տերմին " մատրիցային կենսասինթեզներ «նշանակում է բջջի կարողությունը սինթեզելու պոլիմերային մոլեկուլներ, ինչպիսիք են նուկլեինաթթուներԵվ սկյուռիկներ, հիմնված կաղապարի վրա՝ մատրիցա։ Սա ապահովում է ամենաբարդ կառուցվածքի ճշգրիտ փոխանցումը գոյություն ունեցող մոլեկուլներից նոր սինթեզվածներին:
Մոլեկուլային կենսաբանության հիմնական պոստուլատը
Դեպքերի ճնշող մեծամասնությունում՝ փոխանցում ժառանգական տեղեկատվությունմայր բջիջից մինչև դուստր բջիջ իրականացվում է ԴՆԹ-ի միջոցով (կրկնօրինակում): Բջջի կողմից գենետիկական տեղեկատվության օգտագործման համար անհրաժեշտ է ՌՆԹ, որը ձևավորվում է ԴՆԹ-ի կաղապարի վրա (տրանսկրիպցիա): Ավելին, ՌՆԹ-ն ուղղակիորեն ներգրավված է սպիտակուցի մոլեկուլների սինթեզի բոլոր փուլերում (թարգմանություն), որոնք ապահովում են բջջի կառուցվածքը և գործունեությունը:
Ելնելով վերը նշվածից մոլեկուլային կենսաբանության կենտրոնական դոգմա, ըստ որի՝ գենետիկական տեղեկատվության փոխանցումն իրականացվում է միայն նուկլեինաթթուներից (ԴՆԹ և ՌՆԹ)։ Տեղեկատվության ստացողը կարող է լինել մեկ այլ նուկլեինաթթու (ԴՆԹ կամ ՌՆԹ) և սպիտակուց:
Մոլեկուլային կենտրոնական դոգմակենսաբանություն - բնության մեջ դիտվող գենետիկական տեղեկատվության իրականացումն ընդհանրացնող կանոն. տեղեկատվությունը նուկլեինաթթուներից փոխանցվում է սպիտակուց, բայց ոչ հակառակ ուղղությամբ: Կանոնը ձևակերպվել է Ֆրենսիս Քրիքի կողմից 1958 թվականին և համապատասխանեցվել 1970 թվականին մինչ այդ կուտակված տվյալներին։ Գենետիկական տեղեկատվության փոխանցումը ԴՆԹ-ից ՌՆԹ-ին և ՌՆԹ-ից սպիտակուցին ունիվերսալ է բոլոր բջջային օրգանիզմների համար՝ առանց բացառության և ընկած է մակրոմոլեկուլների կենսասինթեզի հիմքում։ Գենոմի վերարտադրությունը համապատասխանում է ԴՆԹ → ԴՆԹ տեղեկատվական անցմանը: Բնության մեջ կան նաև անցումներ՝ ՌՆԹ → ՌՆԹ և ՌՆԹ → ԴՆԹ (օրինակ՝ որոշ վիրուսների մոտ), ինչպես նաև մոլեկուլից մոլեկուլ փոխանցվող սպիտակուցների կոնֆորմացիայի փոփոխություն։ Տառադարձում և թարգմանություն։Պայմանականորեն, տառադարձման և թարգմանության ողջ գործընթացը կարող է ցուցադրվել հետևյալ գծապատկերում. Տրանսկրիպցիան ԴՆԹ-ում պահվող տեղեկատվության վերարտադրման գործընթացն է միաշղթա մոլեկուլի և ՌՆԹ-ի (սուրհանդակային ՌՆԹ, որը փոխանցում է սպիտակուցի կառուցվածքի մասին տեղեկությունը): բջջային միջուկը դեպի բջջի ցիտոպլազմ՝ ռիբոսոմներ): Այս գործընթացը դրսևորվում է ԴՆԹ-ի կաղապարից մոլեկուլի և ՌՆԹ-ի սինթեզում։ Մոլեկուլը և ՌՆԹ-ն բաղկացած են նուկլեոտիդներից, որոնցից յուրաքանչյուրը ներառում է ֆոսֆորաթթվի մնացորդ, շաքար, ռիբոզ և չորսից մեկը։ ազոտային հիմքեր(A, G, C և U-uracil փոխարեն T-tulin): Սինթեզը և ՌՆԹ-ն հիմնված են կոմպլեմենտարության սկզբունքի վրա, այսինքն. ԴՆԹ-ի մի շղթայում A-ի նկատմամբ Y-ն է և ՌՆԹ-ն, իսկ G-ի դեմ՝ C-ն և ՌՆԹ-ն (տես Նկ. Տրանսկրիպցիան - նախորդ էջում), հետևաբար ՌՆԹ-ն ԴՆԹ-ի կամ դրա որոշակի հատվածի լրացուցիչ պատճենն է։ և պարունակում է տեղեկատվություն, որը կոդավորում է ամինաթթու կամ սպիտակուց: ԴՆԹ-ի և ՌՆԹ-ի յուրաքանչյուր ամինաթթու կոդավորված է 3 նուկլեոտիդների հաջորդականությամբ, այսինքն. - եռյակ, որը կոչվում է կոդոն: Եթե տառադարձման ժամանակ երկու մոլեկուլների ճանաչումը միմյանց կողմից դրսևորվում է միայն փոխլրացման սկզբունքով, ապա թարգմանության մեջ, բացի կոմպլեմենտարությունից (կոդոնի և ՌՆԹ-ի և հակակոդոնի ժամանակավոր համակցություն. ՌՆԹ-ի (փոխադրող ՌՆԹ, որը սպիտակուցի սինթեզի համար անհրաժեշտ ամինաթթուները բերում է սինթեզի վայր՝ ռիբոսոմ - տես Նկ. Տառադարձում) մոլեկուլային ճանաչումը դրսևորվում է ամինաթթու tRNA-ին կոդազ ֆերմենտի միջոցով ամինաթթու կցելու գործընթացում: Փաստն այն է. որ tRNA մոլեկուլը բաղկացած է գլխից, որը ներառում է հակա-AOK եռյակ, որը բաղկացած է երեք նուկլեոտիդների հաջորդականությունից և պոչից, որն ունի որոշակի տեսակի tRNA հակակոսոններ, այնքան պոչերի ձևեր, և յուրաքանչյուր հակակոսոն ունի. tRNA-ում իր պոչի ձևը: Պոչերի քանի ձև կա, այնքան շատ տեսակի կոդազ ֆերմենտի ձևեր, որոնք ամինաթթուները կապում են պոչին, և յուրաքանչյուր կոդազի ձևը համապատասխանում է միայն ձևին: Այսպիսով, tRNA-ն իր հետ տեղեկատվություն է կրում ոչ միայն հակակոզոնի նուկլեոտիդների n հաջորդականությամբ, այլ նաև մոլեկուլի պոչի տեսքով։ Եվ այստեղ տեղեկատվության հիմնական փոխանցումը սպիտակուցում ամինաթթուների հաջորդականության վերարտադրումն է, որը հուշում է սպիտակուցը և ՌՆԹ-ն կոդավորող ֆերմենտը:

| Նախորդ նյութեր. |